
Revenons donc à la biologie. Puisque vous avez précédemment fait allusion aux « champs morphogénétiques », on ne peut pas ne pas penser à la fameuse « loi biogénétique fondamentale » d’Ernst Haeckel, selon laquelle l’ontogenèse récapitulerait la phylogenèse. Bien qu’aujourd’hui on n’ait guère tendance à la mettre beaucoup en relief, cette loi n’en est pas moins considérée par certains comme une preuve que l’évolution a bien eu lieu. Pour sa part, Massimo Piattelli Palmarini, directeur du Centre d’Histoire et de Philosophie de la Science (Florence), a récemment affirmé que, parmi les biologistes, « personne aujourd’hui n’y croit plus » [1], sans doute en exagérant un peu. Que pensez-vous de cette fameuse loi ?
Je pense qu’il serait grand temps de s’en débarrasser une bonne fois pour toutes ! De l’époque où elle fut énoncée pour la première fois à aujourd’hui, en effet, des autorités comme Wilhelm His, Adam Sedgwick, Kart von Baer, Walter Garstang, Gavin de Beer et W.E. Swinton n’ont pas manqué d’en montrer la nature fallacieuse [2].
Il n’y a à coup sûr aucun animal adulte, actuel ou fossile, qu’on puisse dire identique à une quelconque phase embryonnaire d’un autre animal réputé plus évolué. D’autre part, admettre que l’ontogenèse d’un animal donné équivaut à un bref résumé de son histoire évolutive, c’est comme admettre que cette histoire s’est faite par additions successives de nouveaux caractères aux stades adultes des prédécesseurs, et que ces stades adultes, ensuite, ont été, on ne sait trop comment, « miniaturisés » dans les phases embryonnaires de leurs descendants. L’évolution, en somme, serait la cause mécanique du développement ontogénétique !
Bien qu’on ait démontré que les données de Haeckel ont été partiellement falsifiées par lui afin de confirmer l’idée de la récapitulation, et bien que von Baer ait fait clairement remarquer qu’à aucun moment un embryon ne pouvait ressembler à un animal inférieur adulte, mais seulement à l’embryon de ce dernier, pendant très longtemps on a dit, par exemple, que le cœur de l’embryon humain présente d’abord une seule chambre comme chez les Annélides, puis deux comme chez les poissons, puis quatre avec mélange de sang veineux et de sang artériel comme chez les reptiles, et enfin — après la naissance — montre l’oreillette et le ventricule droits bien séparés de l’oreillette et du ventricule gauches, comme chez les oiseaux et les autres mammifères. Un autre exemple était souvent donné : celui des « fentes branchiales », situées entre la tête et le tronc comme chez les poissons. Et puis encore un autre, fourni, lui, par la présence transitoire de reins archaïques : le pronéphros et le mésonéphros.
En réalité, le mélange de sang veineux et de sang artériel dans l’embryon humain doit être mis en relation avec le fait que les poumons ne fonctionnent pas encore, l’oxygénation de l’embryon se faisant au moyen du sang de la mère qui s’écoule à travers le placenta. Quant aux soi-disant « fentes branchiales », elles se ramènent simplement à une série de replis qui ne s’ouvrent jamais et qui ne remplissent donc aucune fonction respiratoire. Les replis sont toujours couverts d’une fine membrane et se transforment en ébauches d’autres structures anatomiques comme la trompe d’Eustache, les thyroïdes, les parathyroïdes et le thymus. Enfin, en ce qui concerne le pronéphros et le mésonéphros, ils ne remplissent aucune fonction excrétoire.
Au cours du développement apparaissent des structures qui peuvent ressembler superficiellement à celles d’autres animaux ; mais dans de nombreux cas, elles n’ont pas du tout les mêmes fonctions. Par ailleurs, différents stades qu’on s’attendrait à rencontrer font totalement défaut, bien souvent, ou se succèdent selon un ordre ne correspondant pas à celui prévu par la thèse de la récapitulation. Les fœtus et les petits du singe, par exemple, présentent des caractères bien plus « humains » que les adultes ; de sorte que, sur la base de la loi de Haeckel, on pourrait très bien en déduire que ce n’est pas l’homme qui descend d’ancêtres de type simiesque, mais, au contraire, que ce sont les singes qui descendent d’ancêtres humains. Et, de fait, les auteurs ne manquent pas qui ont fini par penser de la sorte [3].
Ce qu’il faut surtout souligner, c’est le fait que tous les stades que traverse l’embryon sont nécessaires à l’accomplissement régulier de son développement, de la cellule-œuf fécondée jusqu’à l’organisme adulte ; ces stades doivent donc être vus comme des éléments distincts mais s’intégrant harmonieusement d’une totalité spatio-temporelle unitaire.
Concrètement, ce que vous dites est admis par tous les embryologistes. Quoi qu’il en soit, il ressort clairement du livre Ontogeny and Phylogeny de Stephen Jay Gould [4] que l’auteur est « perplexe » au sujet de l’idée de la récapitulation. D’une part, Gould sait parfaitement que Haeckel a été réfuté ; de l’autre, il croit néanmoins qu’il y a du vrai dans sa position. Comment expliquez-vous ce paradoxe ?
La perplexité de Gould a été critiquée par l’embryologiste suédois Soren Lovtrup [5], certainement l’un des principaux protagonistes des discussions actuelles sur le problème de l’évolution biologique. Lovtrup soutient que le paradoxe dont vous parlez a déjà été résolu depuis très longtemps en distinguant deux façons d’entendre l’idée de la récapitulation : celle de Haeckel et celle de von Baer. La première serait erronée, la seconde juste, et pourrait être formulée ainsi : l’ontogenèse récapitule le cours systématique et le mécanisme de la phylogenèse, ou bien : la succession évolutive des adultes est le produit d’ontogenèses successives. Au cours de son développement, en somme, l’embryon assume les configurations du type, de la classe, de l’ordre, de la famille, du genre et de l’espèce auxquels il appartient ; et puisque — soutient Lovtrup — l’évolution biologique s’est aussi produite de cette façon (du général au particulier), il est clair que l’ontogenèse ne peut pas ne pas récapituler la phylogenèse, fût-ce dans un sens différent de celui soutenu par Haeckel.
Mais je dois dire qu’il me semble illicite d’affirmer — comme le fait Lovtrup — que ce qui était faux, ce n’était pas l’idée de la récapitulation, mais seulement l’interprétation qu’en donnait Haeckel, celle de von Baer étant réputée juste. En réalité, ce dernier ne croyait à aucun type de récapitulation.
Il y a quelques années, Lovtrup a publié dans la Rivista di Biologia un long article en trois parties [6], article apprécié par Sermonti, qui s’est déclaré d’accord pour l’essentiel. Cet article résume le point de vue de Lovtrup sur la question de l’évolution. Il s’agit d’un point de vue qui, d’après ce que j’ai pu voir, s’éloigne beaucoup des thèses courantes. Êtes-vous d’accord avec Lovtrup ?
Non. J’ai beaucoup d’estime pour Lovtrup, mais je ne peux pas accepter le noyau de sa pensée, qui représente à mon avis, sous certains aspects et en dépit de son caractère éclectique, une intéressante reprise de la vieille théorie de l’ologenèse formulée par Daniele Rosa.
En quel sens ?
Pour le comprendre, il faut entrer un peu dans les détails. Lovtrup ne refuse pas les acquis du darwinisme, mais les estime insuffisants pour expliquer le phénomène vivant ; il les complète donc par d’autres acquis, dans le but de parvenir à une théorie vraiment cohérente et complète de l’évolution biologique. Ambitieuse, cette intention a cependant le mérite de mettre de l’ordre dans un domaine si flou et si embrouillé ; elle ne manque pas, en outre, d’une certaine « monumentalité ». J’estime donc que l’œuvre de Lovtrup joue un rôle vraiment important dans le débat sur l’évolution et ne peut pas être ignorée.
Après avoir rappelé que le progrès du simple vers le complexe est un aspect essentiel de la notion d’évolution (« croire à l’évolution et refuser une évolution progressive est une contradiction flagrante » [7], écrit-il), Lovtrup soutient que la « théorie de l’évolution » est en réalité composée de quatre théories, portant respectivement sur : 1) la réalité ; 2) l’histoire ; 3) le mécanisme de formation de nouveaux taxa ; 4) le mécanisme de survie de ces derniers.
La première théorie affirme que la vie sur la Terre s’est développée à travers un processus déterministe d’évolution de forme simples vers des formes toujours plus complexes. Elle serait pratiquement une certitude, puisque la seule documentation paléontologique suffirait — selon Lovtrup — à démontrer que les choses se sont passées ainsi.
La deuxième théorie soutient que l’évolution s’est déroulée dans le temps comme l’imaginait Lamarck et non comme l’imaginait Darwin, c’est-à-dire — en parfait accord avec ce qui se manifeste au cours du développement ontogénétique — du général au particulier, des taxa supérieurs aux taxa inférieurs. Selon Lovtrup, l’espèce n’est donc que le résultat final du processus évolutif ; elle n’a pas le rôle primordial que lui attribuait Darwin (dont l’ouvrage aurait dû s’appeler plus proprement On the Origin of Taxa), et que lui attribuent toujours les néodarwiniens (qu’ils soient partisans de la « théorie synthétique » ou qu’ils se rattachent à l’école des « équilibres ponctués »). L’instrument le plus indiqué pour reconstruire l’histoire de l’évolution serait fourni par la « systématique phylogénétique » de Willi Hennig.
La troisième théorie prétend que les nouveaux taxa doivent se former, non à la suite de petites mutations des gènes structuraux, mais essentiellement sous l’effet de macromutations fortuites intéressant les gènes régulateurs et modifiant le déroulement de l’épigenèse. Lorsque les macromutations concerneront les premières phases de ce déroulement, se produiront les plus grands changements systématiques. Les macromutations concernant les phases postérieures produiront au contraire des changements mineurs. En ce sens, les thèses de Richard Goldschmidt [8], Otto Schindewolf [9] et Albert Dalcq [10] — jusqu’à présent considérées comme de véritables hérésies par les biologistes néodarwiniens — seraient donc réhabilitées et, en partie, vengées.
La quatrième théorie soutient que les taxa nouvellement formés survivent en choisissant les milieux répondant le mieux aux nouveaux caractères qu’ils ont acquis (à la suite de macromutations) et en s’y répandant. Si ces milieux ne sont pas déjà occupés par d’autres taxa, les nouveaux taxa s’y installeront tranquillement ; dans le cas contraire, il y aura compétition, et seuls les dominants, les plus fourbes et les plus chanceux, parviendront à survivre. Ce n’est donc pas la pression de la sélection, mais les forces génétiques et épigénétiques agissant en harmonie avec l’isolement et la compétition interspécifique, qui font naître les nouveaux taxa et assurent leur survie.
Que faut-il penser de cette conception ? Il me semble que le point le plus discutable tient à la façon dont Lovtrup conçoit le processus général de transformation des vivants : des taxa les plus élevés aux plus bas, ou bien des types initiaux très « larges » à des sous-types de plus en plus limités et circonscrits. J’ai le sentiment que ce processus n’équivaut pas du tout à un passage du simple au complexe, mais comporte en revanche une régression, puisqu’au cours du processus sont progressivement exclues des potentialités inhérentes aux types originaires ; donc que cela ne s’accorde pas avec l’idée qu’il faut avoir de l’évolution biologique, d’après Lovtrup lui-même. S’il en est ainsi, comment se sont produits les types originaires : règnes, phyla et classes ? Et si tous les taxa connus ne sont que le résultat de transformations apparues à des phases plus ou moins précoces (= taxa plus ou moins élevés) d’un processus épigénétique préexistant, implicite dans un organisme ancestral, d’où vient le processus épigénétique « totipotentiel » inhérent aux premières formes vivantes apparues sur la Terre ?
En substance, pour Lovtrup l’histoire de la vie s’est déroulée de manière très proche de celle imaginée par le fondateur de l’ologenèse, Daniele Rosa, et par le biogéographe Léon Croizat, auteur d’un gros ouvrage intitulé Panbiogeography et ancien élève de Rosa : une combinaison d’espace, de temps et de forme. À vrai dire, Rosa concevait le processus général de transformation comme déjà programmé dans les premières formes vivantes apparues sur notre planète (exactement comme dans un œuf, où est programmé tout le développement postérieur de l’organisme, avec la différenciation de ce dernier en appareils, organes et tissus) et mû par des facteurs internes (indépendants de ceux du milieu) ; alors que pour Lovtrup, le processus est la conséquence de macromutations totalement casuelles. Mais le tableau du déroulement historique de ce processus est le même.
Comme je l’ai dit, Lovtrup accepte la « systématique phylogénétique » (aujourd’hui appelée aussi « cladistique ») proposée par Hennig, qui s’inspire également, et de façon directe, des contenus de la doctrine de Rosa. Selon cette théorie, le processus transformiste se déroulerait à travers des ramifications dichotomiques successives, dans lesquelles un rameau conserverait les caractères primitifs (ou plésiomorphes) des formes ancestrales, tandis que l’autre donnerait naissance à de nouveaux groupes monophylétiques et se distinguerait par la présence de caractères dérivés (ou apomorphes). Seules la définition et la succession ordonnée des caractères apomorphes pourraient nous donner des informations crédibles sur la façon dont s’est produite la ramification évolutive.
En toute rigueur, il semblerait naturel de supposer que seule la documentation paléontologique puisse nous fournir des indications sûres concernant la distinction des caractères apomorphes et des caractères plésiomorphes. Mais, assez curieusement, ce n’est pas le cas. Les « cladistes », la plupart du temps, ont tendance à ne pas attribuer beaucoup d’importance à cette documentation. Tout se ramène donc à un seul objectif : définir, parmi les possibles arbres phylogénétiques posés comme hypothèses pour justifier l’origine d’un ensemble donné de taxa actuels, le « plus parcimonieux » en éléments et bifurcations. Il s’agit, en résumé, de choisir, parmi les diverses hypothèses généalogiques formulables, la plus simple. Et ce même sans faire référence aux fossiles. C’est la logique du « rasoir d’Occam ».
En dehors de la recherche inductive de l’histoire évolutive — fondée surtout sur les organismes actuels — les « cladistes » soutiennent aussi que les classifications systématiques doivent reposer essentiellement sur les bifurcations résultant de l’arbre généalogique « le plus parcimonieux » auquel a mené l’enquête. Mais puisqu’il arrive parfois qu’il y ait plus d’une solution parcimonieuse, et puisqu’il n’est pas dit que tous les auteurs seront d’accord sur la signification effective (apomorphique ou plésiomorphique) à attribuer aux caractères, il s’ensuit que la systématique continuera de s’appuyer sur une base très instable et non exempte de subjectivisme.
Comme bon nombre d’autres « cladistes », Lovtrup affirme que les « classifications phylogénétiques » doivent avant tout reposer sur les organismes actuels. Les fossiles, en effet, ne fourniraient que des informations éparses et fragmentaires ; ils devraient donc être, selon lui, étudiés et classés à part. Sur ce point, très peu de paléontologues seront d’accord avec Lovtrup. Nous pouvons tous être d’accord pour dire, par exemple, que le Tyrannosaurus était plus proche des lézards et des crocodiles que des loups et des jaguars actuels, et qu’il doit donc être classé avec les reptiles plutôt qu’avec les mammifères. Mais, par là même, ne l’avons-nous pas déjà classé avec les organismes actuels ?
Lovtrup n’est pas non plus très convaincant au sujet du mécanisme du processus de transformation. Ses « macromutations » ne correspondent ni aux « mutations systémiques » de Goldschmidt, ni aux « ontomutations » de Dalcq (que personne n’a jamais pu observer et étudier expérimentalement), et ne sont pas de nature qualitativement différente de celles étudiées par les généticiens. Leur seule particularité semble être de se produire surtout au niveau des gènes régulateurs, des chromosomes (avec duplications, inversions, etc.) et du génome en général (à travers polyploïdie, aneuploïdie, etc.), pour s’amplifier ensuite au cours des processus épigénétiques. Jusqu’ici, parfait. Mais le fait est que l’embryologiste suédois ne croit pas au « dogme central de la biologie », selon lequel l’ADN est la cause efficiente primaire de toute structure biologique. « Personne, je pense, ne s’attendrait à ce que le noyau cellulaire, placé dans un milieu approprié, s’entourât d’un cytoplasme et d’une membrane pour former ainsi une cellule » [11] écrit Lovtrup. Au contraire, le développement épigénétique, loin d’être contrôlé par le seul génome, résulterait de l’interaction constante entre ce dernier et le cytoplasme, en fonction aussi des influences du milieu. Il est même possible de démontrer que les premières phases de ce développement — les plus importantes, précisément, sur le plan morphogénétique — sont aussi les moins étroitement dépendantes de l’activité du génome.
Or, si tout cela est vrai, pourquoi le processus de transformation naturelle devrait-il survenir uniquement par l’intermédiaire de macromutations apparues dans le génome ? Il me semble que Sermonti [12] a parfaitement raison de critiquer cette incohérence dans la conception de Lovtrup.
Mais comment peut-on nier avec certitude que des mutations au niveau des gènes régulateurs, des chromosomes et du génome conduisent à la formation de nouveaux taxa ?
Et comment peut-on l’affirmer avec certitude ?
Si vous répondez ainsi, nous nous retrouvons dans un cercle vicieux.
Pas du tout. Ne croyez pas que je veuille à tout prix me faire l’« avocat du diable ». À ce sujet, dire que nous sommes dans l’incertitude la plus totale, c’est tout simplement dire la vérité. L’éminent généticien Richard Lewontin l’a lui-même reconnu dans son fameux ouvrage The Genetic Basis of the Evolutionary Change : « Nous ne savons pratiquement rien sur les changements qui se vérifient dans la formation des espèces » [13]. Il me paraît extrêmement significatif que Lewontin ait placé en exergue de son livre les deux premiers tercets de la Divine Comédie : « Nel mezzo del cammin di nostra vita / mi ritrovai per una selva oscura, / che la diritta via era smarrita. / Ah quanto a dir qual era è cosa dura / esta selva selvaggia ed aspra e forte / che nel pensier rinnova la paura ! » [14].
Évidemment, la « forêt obscure » à laquelle pensait Lewontin, c’est le problème posé par la terrible variabilité et instabilité des vivants, variabilité et instabilité qu’il rencontra pour la première fois, il y a vingt ans environ, au niveau génétique et moléculaire, et qui n’étaient pas du tout prévues par la théorie synthétique néodarwinienne.
Dans l’intérêt du lecteur, il me semble nécessaire que vous présentiez, au moins dans ses grandes lignes, cette théorie.
Entendu. Mais auparavant, je voudrais bien souligner ceci : s’il est vrai, incontestablement, que la biologie a été profondément influencée, de 1859 à nos jours, par la théorie darwinienne, il est tout aussi vrai que celle-ci n’a pu exercer une sorte de monopole sur la pensée théorique en biologie qu’après que l’Europe soit devenue culturellement dépendante des puissances victorieuses dans la deuxième Guerre mondiale. Avant celle-ci, en effet, le succès du darwinisme n’avait jamais été complet, car il avait toujours été limité par l’opposition des néolamarckiens (Eimer, Rabaud, Caullery, McDougall, Cuénot, etc.), des vitalistes (Driesch, Berg, Rignano, Rosa, etc.) et de tout le courant des biologistes « holistes » qui, prenant leurs distances par rapport à l’évolutionnisme — darwinien ou lamarckien — et au vitalisme, avaient résolument emprunté la voie tracée par la idealistische Morphologie de Goethe et par l’organicisme aristotélicien (von Uexküll, Troll, Dacqué, von Bertalanffy, Smuts, Haldane, Russel, Vialleton, Raffaele, Pasquini, Canella, etc.).
Après la seconde Guerre mondiale, il était vraiment impossible que les thèses des biologistes holistes pussent être acceptées par la mentalité des vainqueurs américains et soviétiques. Tout naturellement, les premiers se reconnaissaient dans l’idée de « sélection des meilleurs » résultant de la libre compétition, et les seconds dans l’idée de la puissance transformatrice de l’environnement physique et social — l’une et l’autre vues comme sources de tout progrès —, idées qui sont absolument inhérentes à la vision matérialiste de Darwin. Le développement de la biologie holiste fut donc entravé, partout dans le monde, par l’entente « syndicale » de ceux qui posèrent les bases de ce qu’on appelle la « théorie synthétique » néodarwinienne, au sein de laquelle la sélection naturelle se combine avec la génétique de Mendel et le mutationnisme de De Vries. Les ouvrages d’évolutionnistes réputés mais étrangers au « syndicat » — comme Goldschmidt, Schindewolf et Colosi — furent discrédités et présentés comme des produits de la pure fantaisie.
Les thèses affirmant que tout ne pouvait pas être ramené aux seules lois du hasard et de la nécessité furent écartées comme antiscientifiques, hâtivement mises à l’index ou froidement ignorées. On en arriva même à proclamer que la théorie synthétique, ou « synthèse moderne », non seulement avait liquidé tout finalisme (« concept inutile, né de la pensée grecque, qui était anthropomorphique de manière sincère et primitive », dira Simpson), mais avait aussi contraint à la révision de tous les concepts éthiques et métaphysiques, permettant ainsi de jeter à la mer toute la pensée « typologique » ou « essentialiste » (laquelle, dira Mayr, « prétend que le monde changeant des apparences est fondé sur d’immuables essences sous-jacentes, et que tous les membres d’une classe représentent la même essence »).
La théorie synthétique repose essentiellement sur trois ouvrages, parus aux Etats-Unis dans la première moitié des années quarante et se complétant très bien entre eux : Genetics and the Origin of Species de Theodosius Dobzhansky (un généticien russe qui avait émigré en Amérique), Systematics and the Origin of Species d’Ernst Mayr (un naturaliste allemand qui avait lui aussi émigré), et Tempo and Mode in Evolution du paléontologue George Gaylord Simpson.
Dobzhansky soutenait que les variations génétiques responsables du processus évolutif sont quantitativement très réduites et transmissibles selon les lois de Mendel. Ces variations ne seraient que des formes différentes (ou allèles), produites à la suite de mutations, de gènes gouvernant certains caractères biologiques. Dans une population, les individus peuvent porter tel ou tel de ces allèles, et présenter par conséquent l’une ou l’autre des constitutions de l’organisme dictées par les divers gènes allèles. Certaines constitutions assureraient aux individus une meilleure capacité d’adaptation au milieu, si bien qu’ils vivraient plus longtemps et se reproduiraient plus que leurs concurrents. Il s’ensuivrait que dans les générations suivantes les gènes allèles responsables des constitutions mieux adaptées deviendraient de plus en plus fréquents dans la population, pour finir par être exclusifs des autres. Si ce remplacement d’allèles par des allèles plus adaptés concerne un nombre élevé de gènes, la population finira par acquérir une constitution génétique très différente de celle de la population initiale : une nouvelle espèce naîtra.
À son tour, Mayr introduisait trois notions importantes : le concept « biologique » d’espèce, la variation géographique de l’espèce et la spéciation allopatrique. Ces trois notions permettraient de comprendre comment devait se produire, vraisemblablement, la formation des nouvelles espèces dans la nature. Le concept biologique d’espèce impliquait que celle-ci est un ensemble de populations naturelles formant une communauté reproductrice unique et reproductivement isolée d’autres communautés semblables. La variation géographique de l’espèce renvoie au fait que les espèces sont distribuées dans des territoires souvent très distants les uns des autres, si bien que les conditions environnementales ne sont pas semblables pour toutes les populations faisant partie de la même espèce. Il s’ensuit que les populations très éloignées entre elles présentent des caractères adaptatifs différents (par exemple la taille des animaux à sang chaud d’une même espèce tend à être plus grande dans les pays froids et plus petite dans les pays chauds, en rapport avec la régulation thermique), de sorte qu’elles peuvent former des races géographiques au sein des mêmes espèces. Lorsque certaines populations seront géographiquement séparées de la zone principale de distribution de l’espèce (comme par exemple dans une île), leurs caractères biologiques pourront diverger assez fortement de ceux des populations de l’aire principale, au point de ne plus pouvoir autoriser des croisements avec ces derniers et de ne plus pouvoir donner de descendants fertiles. La population isolée de l’ensemble de populations dont elle faisait partie deviendra alors une communauté reproductrice autonome, c’est-à-dire une espèce nouvelle.
Simpson, enfin, s’efforçait de rattacher la paléontologie à la génétique des populations de Dobzhansky et de Mayr, en soutenant que l’évolution consistait dans l’accumulation graduelle de petites variations au sein des populations. Il mit au point des méthodes de quantification des données paléontologiques (comme par exemple la mesure des taux d’évolution dans les différentes lignes phylétiques) pour établir si les modes d’évolution observés dans la documentation fossile sont compatibles ou non avec le modèle proposé par la génétique des populations ; et il arriva à la conclusion qu’ils l’étaient effectivement. À ceux qui objectaient que l’évolution se semblait pas s’être déroulée fortuitement, mais selon des lignes directrices nettement orientées (ou par orthogenèse), Simpson répondait que ces orientations linéaires — quand elles existaient réellement (et dans des cas comme celui des Équidés elles n’existaient certainement pas) — ne résultaient pas d’une tendance finalisée immanente dans les organismes vivants, mais du maintien dans la même direction de tendances favorisées par la sélection naturelle. Il n’y aurait donc pas orthogenèse, mais seulement orthosélection.
La construction « synthétique », indubitablement très simple et fascinante, fut au faîte de sa réussite au moment même où l’on célébra le centième anniversaire de la parution de Sur l’origine des espèces, donc en 1959. Mais elle n’avait jamais prouvé qu’elle était capable de résister à une critique vraiment approfondie et sans passion, et déjà vers le milieu des année soixante ses propres partisans commencèrent à découvrir un nombre croissant de faits qui n’étaient pas du tout faciles à expliquer dans le cadre théorique dont ils se servaient. En 1966 Lewontin, élève de Dobzhansky, découvrit avec J. H. Hubby que les organismes ont, en général, un taux d’hétérozygosie [15] plus élevé que ce qu’on croyait (évalué ensuite à environ 17 % pour les Plantes, 13 % pour les Invertébrés et 6 % pour les Vertébrés), et aussi, assez fréquemment, des allèles multiples (constitués, non de deux formes seulement d’un gène, mais parfois de plusieurs douzaines de formes alternatives).
Dès lors, il est clair qu’il est devenu plus que légitime de se demander si la sélection, elle seule, peut rendre compte de ce déconcertant polymorphisme biologique. Et nombreux ont été les généticiens qui ont estimé qu’il fallait répondre négativement. Il suffira de citer Motoo Kimura et toute son école (Jack L. King, Thomas H. Jukes, Tomoko Ohta, James F. Crow, Masatoshi Nei, etc.) [16] pour eux, la plupart des changements qui ont lieu au niveau génético-moléculaire — et donc, évidemment, le processus même d’évolution biologique — ne peuvent pas être vus comme produits par l’action de la sélection naturelle (ils seraient donc sélectivement neutres, car cette dernière n’aurait pas prise sur eux) et seraient uniquement sujets à l’action de la « dérive génétique » (genetic drift). Il s’agirait en somme de simples allèles mutants qui, une fois formés, se répandraient dans la population, avec une fréquence fluctuant dans le temps, avant de s’éteindre. Et puisque tout cela serait soumis au hasard, on peut dire que la théorie « neutraliste » de l’évolution moléculaire est encore plus réductrice que la théorie synthétique, qui avait au moins le mérite d’associer la nécessité au hasard.
Mais de temps en temps, à des intervalles dont on estime qu’ils peuvent être mesurés avec une certaine précision, un allèle mutant se fixerait dans la population, si bien que la vitesse d’évolution au niveau moléculaire serait relativement constante pour tous les vivants, et indépendante du temps de génération (la fréquence avec laquelle un organisme se reproduit), des fluctuations environnementales et des dimensions des populations. On admet que cette vitesse peut être évaluée par l’analyse des molécules protéiques et d’ADN (en comptant en elles le nombre d’acides aminés et de nucléotides différents) et par des informations chronologiques sûres fournies par la géologie stratigraphique. L’hémoglobine de l’homme, par exemple, diffère de celle du rat d’environ 20 acides aminés, de celle du triton d’environ 60 acides aminés et de celle du requin d’environ 80 acides aminés. En connaissant le temps approximatif d’apparition des requins dans la succession stratigraphique et paléontologique, on estime que la vitesse moyenne d’évolution moléculaire de l’hémoglobine correspond, plus ou moins, à un remplacement d’acide aminé tous les 7 millions d’années.
Vincent Sarich, Alan Wilson et d’autres considèrent la vitesse d’évolution moléculaire comme un moyen infaillible pour dater les bifurcations qui, selon eux, ont eu lieu entre les différents groupes systématiques : un moyen tout aussi précis que celui fourni par les méthodes de datation absolue fondées sur l’émission des éléments radioactifs. Ainsi, pour donner un exemple, ils estiment que la ligne humaine et celle des Pongidés (gorille et chimpanzé) se sont séparées il y a seulement 4,5-5 millions d’années, et non pas il y a 20-25 millions d’années comme on l’a généralement affirmé jusqu’à maintenant. Et cela serait précisément démontré par la très haute affinité existant au niveau biochimique entre ces trois formes [17].
Et votre point de vue, quel est-il ?
Je ne crois pas à ces « bifurcations évolutives ». Je suis convaincu que, plus deux organismes sont éloignés entre eux dans l’échelle systématique, plus leurs différences aux niveaux morphologique et biochimique doivent se révéler fortes. Mais la distance systématique entre deux ou plusieurs formes est une chose, la distance entre leurs temps d’apparition dans la succession stratigraphique et fossilifère en est une autre. Dans ce deuxième cas, à mon avis, seule la paléontologie peut avoir le dernier mot.
Cela étant, il faut reconnaître que les argumentations des « neutralistes » sont discutables. On a fait remarquer, par exemple, que l’existence d’espèces entièrement homozygosiques (comme le moucheron du fruit Drosophila odontophallus et le rat-kangourou Dipodomys panamintinus) n’est pas expliquée par la théorie des mutations neutres. Si la grande majorité des polymorphismes dans une espèce est due à des allèles mutants apparus par pur hasard et non parce que préservés par la sélection naturelle, l’absence de polymorphisme indiquerait, en effet, que des mutations ne se produisent pas dans ces espèces. Pourquoi ?
Mais il y a plus. R.C. Richmond [18] a souligné qu’en dehors des incertitudes des datations géologiques, les vitesses d’évolution moléculaire estimées sur des protéines différentes ne sont généralement pas les mêmes : ce qui contredit l’uniformité de cette vitesse, pour tous les vivants, affirmée par les « neutralistes ». En outre, c’est l’un des plus connus parmi ces derniers, Nei, qui reconnaît que la théorie des mutations neutres est fortement handicapée du fait qu’elle soutient que le taux de mutations neutres reste constant année par année, plutôt que génération après génération. « Si cette affirmation est fausse, écrit Nei, la théorie de la mutation neutre sera sérieusement compromise. Nous avons présenté (…) certaines évidences en renfort de cette affirmation, mais le taux de mutations neutres reste largement inconnu » [19]. L’affirmation en question semble en fait sans fondement. Georges Pasteur écrit de son côté : « Les biochimistes qui comparent les chaînes d’ADN aboutissent à des résultats diamétralement opposés à ceux qu’obtiennent les chercheurs qui comparent les chaînes protéiques : l’évolution des molécules d’ADN non répété, indépendante du temps absolu (calculé par les géologues, N. D. L. A.), dépend de la durée des générations ! » [20].
On voit donc que la théorie des mutations neutres n’est pas exempte de défauts. Lewontin lui-même ne l’accepte pas intégralement. Il se contente, pour l’essentiel, de mettre en évidence le fait que la génétique des populations classique, fondée sur la « théorie synthétique » néodarwinienne, est incapable de résoudre les problèmes posés par le polymorphisme élevé présent au niveau génético-moléculaire, et met soigneusement en garde les biologistes contre les prétentions de cette théorie, qui voudrait être la vraie et seule explication possible des mécanismes évolutifs, en rappelant les obstacles auxquels ont conduit les modèles mathématiques fondés sur elle.
Lewontin pense que la théorie mathématique de la génétique des populations risque de se réduire à un pur « exercice de logique formelle, sans aucun lien avec le monde réel », non seulement parce que le changement de comportement, l’adaptation et la pression sélective sont difficiles à quantifier et à mesurer, mais surtout parce qu’elle considère les gènes isolément et comme s’ils étaient des choses indépendantes entre elles, au lieu de les voir tels qu’ils sont réellement, c’est-à-dire comme des unités en interaction au sein du génome.
Le fait incontestable de l’interaction génétique ressort du reste tout particulièrement de l’absence de corrélation entre structures corporelles homologues et sites géniques identiques, ainsi que de l’existence des transposons, ou « gènes errants », découverts par le Prix Nobel Barbara McClintock. Au sujet du premier cas, en effet, il est faux de dire — comme l’a également montré Alister Hardy [21] — que l’aile d’un oiseau, le membre antérieur d’une chauve-souris ou celui d’une grenouille, pour ne donner que ces exemples, sont contrôlés par des gènes situés dans les mêmes positions chromosomiques. En réalité, les structures biologiques homologues sont gouvernées, moins par des gènes particuliers, que par le résultat du dynamisme et du réarrangement continus de tout le matériel génétique. Et ce dynamisme continu n’est pas simplement l’effet d’un certain nombre de gènes qui opèrent indépendamment les uns des autres, il est le reflet de leur interaction permanente. Quant aux transposons (dissocation genes, ou Ds), ils « sautent » d’une position à l’autre du chromosome d’après des instructions précises d’autres gènes (activator genes, ou Ac), si bien que leur comportement n’est pas fortuit, mais obéit à de précises normes de régulation.
Alors, dites-moi maintenant : tout cela ne vous rappelle-t-il pas quelque chose ?
Nous sommes revenus, me semble-t-il, au concept holiste d’interaction entre des éléments différents d’un système organique unitaire.
Précisément. Et c’est là le paradoxe de la biologie au XXème siècle. Jusqu’à nos jours, la plupart des biologistes, fidèles à la mentalité réductionniste de la « théorie synthétique », se sont engagés dans l’étude analytique de plus en plus circonscrite des structures héréditaires : nucléaires, cytoplasmiques et macromoléculaires en général. On espérait en effet, de cette façon, arriver à établir ce qu’il y avait de réellement commun et de réellement distinctif — à un niveau de causes aussi élémentaires que possible — entre les innombrables formes organiques existant dans la nature, de l’algue au chêne et de l’amibe à la baleine. Mais tout s’est exactement passé comme en physique subatomique : au terme de ces recherches toujours plus minutieuses, est apparu un dynamisme déconcertant de processus de démantèlement, de reconstruction, d’autoduplication, d’ouvertures et de fermetures « à glissière », de dissociations, de sauts et de recombinaisons, de production de « magasins d’information non utilisée » (les pseudogènes), avec au fond du décor tout cet ADN « répétitif », redondant (carrément « parasite ») et neutre face à des caractérisations systématiques, puisque réputé incapable de transmettre des informations. Et si l’on songe que tout cet incessant dynamisme constitue précisément la matrice de formes ou entités ordonnées qui peuvent être ramenées — dans leur ensemble, et malgré leur plasticité et leur variabilité extrêmes — à un nombre défini de plans de structure fondamentaux, lesquels sont restés stables et inchangés pendant des millions et même des centaines de millions d’années ; si l’on songe à tout cela, alors se pose de nouveau le très vieux problème représenté par la complémentarité harmonieuse entre forme et matière, stabilité et devenir, ordre et désordre, problème plus que jamais actuel en dépit de plus d’un siècle de darwinisme et de plus d’un demi-siècle de génétique « micrométrique ».
J’aimerais que nous nous arrêtions un peu sur le problème de la spéciation, qui me semble vraiment un problème-clé. Nous pouvons admettre, avec Lewontin, que nous ne savons pratiquement rien à ce sujet ; mais toutes les « certitudes » anciennes (je me souviens de ce que j’ai appris à l’université…) se sont-elles vraiment révélées sans fondement ?
Je crains bien que oui, et je crois qu’à ce sujet la seule chose sûre, c’est encore une fois l’insuffisance de l’explication « synthétique ». Outre Lewontin, Hampton L. Carson [22] a lui aussi souligné que la génétique des populations classique, si elle pouvait certainement servir décrire des changements de fréquence de gènes dans les populations, n’en était pas pour autant capable de fournir une explication adéquate de la genèse des espèces.
En résumé, les biologistes actuels sont d’accord sur le fait que le taux de mutation, tout en étant responsable — en définitive — de toute la variabilité des populations, ne joue pourtant pas un rôle important dans les processus de spéciation. Si l’on repousse la théorie des mutations neutres, les adaptations et les pressions sélectives apparaissent alors incroyablement plus raffinées et plus variables que ce qu’on avait supposé jusqu’à maintenant. En dépit des nombreuses controverses à ce sujet, le rôle de la genetic drift (phénomène de la fluctuation de fréquence des gènes) dans la variabilité à l’intérieur d’une population et dans la différenciation entre populations, est encore obscur. L’importance des migrations dans les processus de spéciation est encore très difficile à évaluer, et les nombreux modèles mathématiques qui ont été faits à ce sujet, ont rarement été testés par des observations précises dans la nature. Même la valeur de la panmixie (« participation à la reproduction de tous les individus appartenant à une même population dans le plus grand hasard et en l’absence de toute sélection », selon la définition de Grassé) a été mise en doute.
Tout cela fait que l’attention des biologistes s’est tournée surtout, ces derniers temps, vers l’étude de la structure des populations. Il est évident, en effet, que celle-ci doit influencer fortement la probabilité d’accouplements entre individus, même si nous ne savons pas grand-chose sur les relations entre cette structure et la variabilité génétique [23].
Malgré toutes ces incertitudes, une chose ressort clairement de l’énorme quantité d’études effectuées : le caractère insuffisant des modèles de spéciation invoqués par la « théorie synthétique », à savoir le modèle sympatrique et le modèle allopatrique.
En quoi diffèrent exactement ces deux modèles ?
C’est plutôt simple. Le modèle sympatrique suppose que les individus d’une espèce, en s’adaptant à une niche écologique particulière existant au sein de la zone de distribution de l’espèce elle-même, restent liés à cette niche, ne s’accouplent qu’entre eux et refusent tout contact avec d’autres individus « de passage ». Avec le temps, l’isolement de la niche déterminerait aussi l’isolement génétique proprement dit. Le modèle sympatrique, accepté par Darwin et R. A. Fisher, repris récemment par John Maynard-Smith, a été très critiqué, surtout par Ernst Mayr [24]. Le zoologiste américain Guy Bush [25] ne l’a jugé valable que pour les espèces parasites ; mais, dans ce cas, l’évolution de l’espèce surviendrait de façon brusque, non progressive, ce qui ne correspond donc pas à la théorie darwinienne.
Selon le modèle allopatrique, la spéciation se produirait dans des populations assez restreintes, restées isolées dans des zones périphériques par rapport à l’aire de distribution de l’espèce-mère, à cause de la présence de barrières géographiques (fleuves, montagnes, conditions particulières de végétation, etc.), éthologiques ou autres. Ces populations auraient été soumises à des régimes de forte instabilité environnementale qui auraient provoqué une élévation brutale du taux de mutation, mutations sur lesquelles aurait immédiatement opéré la sélection naturelle. Les isolats périphériques auraient donc connu des conditions leur permettant d’évoluer beaucoup plus rapidement que les représentants de l’espèce-mère, d’atteindre rapidement l’isolement reproductif de cette dernière et de devenir ensuite des espèces distinctes.
Considéré jusqu’à date récente comme le seul vrai, le modèle allopatrique a été de plus en plus contesté au cours des dix dernières années. Il semble en effet que les petites populations soient sujettes à des taux de mutation moins élevés que les grandes. Surtout, Carson a fait remarquer que la spéciation ne peut se produire qu’à la suite d’événements catastrophiques et brusques, probablement entraînés par des cycles démographiques comprenant un phénomène d’expansion rapide dans des niches écologiques disponibles, immédiatement suivi par une baisse progressive de vitalité de la population concernée. Enfin, le fondateur de la cytogénétique, l’Australien Michael J. D. White [26], a porté un dernier coup dur au modèle de spéciation allopatrique en proposant un nouveau modèle — appelé par lui stasipatrique —, qui explique que la spéciation surviendrait essentiellement à la suite de réarrangements au niveau chromosomique, et ce même en l’absence de barrières géographiques.
Cela semble donner raison à Lovtrup, du moins pour ce qui concerne l’origine des seules espèces.
Vous avez raison de dire : semble. Le fait est que, jusqu’à présent en tout cas, on n’a pas été capable de préciser la signification exacte de ces mutations et réorganisations au niveau chromosomique, dans le cadre d’une possible spéciation ou, plus généralement, de la formation de nouveaux taxa. Même si l’on admet qu’il y a une relation étroite entre différences chromosomiques et spéciation, on ne sait pas très bien si ce sont les premières qui déterminent la seconde, ou l’inverse. En d’autres termes, nous ne sommes pas certains que des mutations chromosomiques produisent effectivement des transformations évolutives au niveau morphologique, car s’il est vrai qu’en général les espèces diffèrent entre elles par le nombre et la structure des chromosomes, il est tout aussi vrai qu’il y a de très nombreux exemples d’espèces qui, sans présenter de différences visibles au niveau chromosomique, sont cependant morphologiquement différentes ; et vice versa. Il n’y a pas non plus de corrélation nécessaire entre l’interfécondité et la ressemblance morphologique, puisqu’on connaît des espèces qu’il est impossible de distinguer sur le plan morphologique, mais qui sont isolées sous l’angle reproductif, et des espèces morphologiquement distinctes et géographiquement séparées, mais interfécondes.
Le problème de la spéciation renvoie automatiquement à celui de la distribution géographique des vivants. Ce dernier problème, aujourd’hui, est-il envisagé de la façon dont le voyait Darwin, ou bien y a-t-il eu des nouveautés ?
Je dirais qu’à ce sujet également les biologistes sont en train de passer d’explications de type « hasard et nécessité » à des explications de type holiste.
Pour Darwin, Wallace, Matthew et leurs modernes épigones (Simpson, Mayr, Haldane, Darlington et bien d’autres), les choses étaient relativement simples. Ils croyaient que les données fondamentales de la distribution géographique des vivants pouvaient être expliquées simplement en recourant aux « centres d’origine », avec des migrations postérieures depuis ces centres, migrations dues pour une large part à une dispersion fortuite, entraînant la production, la multiplication et la modification de nouvelles formes. Jusqu’à aujourd’hui, par conséquent, la biogéographie s’est occupée avant tout de centres d’origine, de moyens et de routes de dispersion, de clines (gradients morphologiques liés à des facteurs géographiques), d’évolution darwinienne et d’analyses de composants biotopiques en termes d’éléments colonisateurs.
Mais grâce à l’œuvre monumentale — souvent marquée par un ton polémique — d’un grand chercheur solitaire récemment disparu, Léon Croizat [27], on a pu constater que l’interprétation darwinienne des phénomènes biogéographiques était très insatisfaisante. La distribution de très nombreuses variétés d’organismes à la surface de la Terre n’est pas du tout fortuite, si bien qu’il faut l’étudier d’un point de vue synthétique, comprenant la recherche des vicissitudes géologiques traversées par les territoires où les différentes espèces vivantes sont aujourd’hui réparties.
Pour Croizat, chaque taxon est caractérisé par une certaine « trace » (track) à la surface de notre planète. Une trace, c’est, essentiellement, une relation géographique entre les membres d’un taxon, qu’ils soient distribués uniformément ou en clusters isolés. Or, lorsque la trace d’un taxon donné, par exemple une espèce, se superpose dans une mesure plus ou moins grande aux traces d’autres espèces, toutes tendant à se concentrer dans une même zone, chaque trace peut être vue comme une partie intégrante d’un système que Croizat appelle « trace généralisée » et qui, en lui-même, représente un véritable biotope. Ainsi, chaque trace biologique individuelle ne pourra être correctement comprise et expliquée que par référence à la trace généralisée dont elle fait partie.
Le paradigme suivi par Croizat est donc clairement holiste ou organiciste. En l’adoptant, on ne perd plus de temps à spéculer sur les causes de la distribution biogéographique, ce que font par contre les darwiniens lorsqu’ils se consacrent à la recherche des « centres d’origine » d’où seraient parties les migrations colonisatrices d’autres territoires. Ces « centres » ressemblent extraordinairement aux fameux « chaînons de liaison » des spéculations d’ordre phylogénétique, dont l’existence est toujours admise bien qu’on ne soit jamais parvenu à la démontrer. Au contraire, il s’agit ici de travailler, de recueillir régulièrement des données concrètes de nature biogéographique et géologique se rapportant respectivement aux taxa étudier et à leurs traces, ainsi qu’au plus grand nombre possible de taxa dont la distribution géographique coïncide plus ou moins avec celle des premiers. Et ceci dans le but, précisément, de parvenir à reconstituer les traces généralisées dont les taxa concernés sont partie intégrante.
La méthode utilisée par Croizat pour retrouver les traces généralisées s’appelle méthode panbiogéographique, ou méthode de la vicariance. Elle est fondée dans une large mesure sur l’étude approfondie des changements géomorphologiques qui ont concerné la zone des taxa, contribuant ainsi à provoquer leur subdivision en taxa postérieurs de niveau plus bas. Par exemple, si dans la zone ou trace d’une espèce donnée s’est formée, au cours du temps, une chaîne montagneuse qui l’a coupée en deux, les représentants de l’espèce originelle restés isolés en-deçà et au-delà de la chaîne montagneuse auront atteint, entre-temps, l’isolement génétique et seront devenus deux espèces distinctes. De la sorte, à travers une combinaison d’événements géomorphologiques, climatologiques, et de spéciation allopatrique, nous aurons une biogéographie de la vicariance, où les divisions successives d’un taxon donné prendront la place de ce que sont, dans la théorie darwinienne des centres d’origine, les arrivées ou les multiples colonisations successives, ou encore les migrations séquentielles de taxa venant d’ailleurs (figure 11).
-
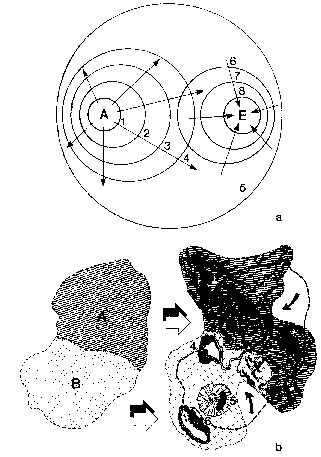
Figure 11
Théories biogéographiques du centre de dispersion (Simpson, Mayr, Darlington, etc.) et de la vicariance (Croizat).
Selon la théorie du centre de dispersion (a), chaque nouveau taxon se forme dans une localité géographique bien déterminée (A, forme autochtone), se répand progressivement par migration active ou passive (phases 1-5) et, après avoir atteint son expansion maximale dans sa zone de distribution (phase 5), finit par se retirer dans des territoires de plus en plus réduits et circonscrits (phases 6-8), souvent même très éloignés du centre d’origine (E, forme endémique).
Selon la théorie de la vicariance (b), en revanche, l’apparition de nouveaux taxa dans une aire géographique bien précise ne signifie pas qu’ils viennent d’ailleurs, mais résulte simplement de la modification in situ de parties de populations préexistantes, à la suite de leur adaptation à de nouvelles conditions géographiques et topographiques (comme la formation de chaînes de montagnes, de lacs, fleuves, volcans, etc.) et de leur isolation du reste des populations-mères (voir par ex. les taxa 1-3 et 4-6, qui se sont formés par ségrégation des populations vicariantes originelles A et B, respectivement). Tout en étant fondamentalement immobiliste, la théorie de la vicariance n’exclut pas une certaine mobilité, qui reste cependant limitée, généralement, aux territoires entourant les zones originelles de distribution, mobilité qui est indiquée sur la figure par les flèches.
La méthode de la vicariance mène à des inférences sur l’histoire de la Terre qui peuvent être vérifiées par les méthodes géophysiques modernes. Inversement, des hypothèses géophysiques peuvent être corroborées par la méthode panbiogéographique. Par exemple, l’idée que l’archipel néo-zélandais faisait partie d’un vaste continent méridional plus ancien (comprenant l’Amérique du Sud, l’Afrique du Sud et l’Australie, et appelé le Gondwana) peut être mise en rapport avec le fait que les traces généralisées intercontinentales des formes vivantes dans l’hémisphère sud incluent ou non les traces biologiques de cet archipel. Si un taxon donné ou un groupe de taxa néo-zélandais faisai(en)t partie de cet ancien continent disparu, cela doit en somme être prouvé par le fait qu’il(s) coïncide(nt) ou non avec une trace généralisée « gondwanienne ».
En résumé, pour Croizat l’histoire des ensembles biotopiques du monde entier et, jusqu’à une forte extension, celle des différents taxa organiques, coïncident avec l’histoire des terres émergées et des océans. L’histoire de la Terre et celle de la vie sont donc considérées comme des développements complémentaires, incluant des périodes d’immobilité (où les espèces, en l’absence de barrières géographiques, se déplacent librement) et des périodes d’immobilité (où la formation de races et les événements spéciationnels apparaissent comme le résultat de l’établissement de barrières géographiques qui fragmentent les aires de distribution).
Deux biologistes nord-américains, Gareth Nelson et Donn Eric Rosen [28], se sont récemment approprié ces idées, jusqu’à présent délibérément ignorées parce qu’incompatibles avec les thèses d’inspiration darwinienne. Ils les ont fait connaître sur le plan international, après les avoir étroitement associées à la théorie phylogénétique et systématique de Hennig. Il me paraît difficile de dire si cela revient à compléter et à perfectionner les idées de Croizat, ou si cela débouche sur leur reductio ad absurdum à la philosophie « cladistique ». Quoi qu’il en soit, selon ces deux auteurs, le processus évolutif des vivants aurait consisté essentiellement dans la subdivision, pendant le temps géologique, de types biologiques généralisés en sous-types de plus en plus circonscrits, tant sous l’angle systématique que sous l’angle géographique. Exactement comme dans le développement épigénétique individuel.
Il me semble qu’il y a, là aussi, des convergences avec ce que soutient Lovtrup.
Incontestablement. Et Lovtrup, pour sa part, cite volontiers, de temps en temps, des phrases de Croizat. Mais il affirme admettre, en plus de la vicariance géographique, la dispersion fortuite à partir de centres d’origine, et ajoute qu’il ne comprend pas bien les opinions de Croizat au sujet du mécanisme du processus évolutif.
En quoi consisterait, selon Croizat, ce mécanisme ?
Pour être franc, je dois dire que moi aussi je n’ai pas trouvé Croizat très clair à ce sujet. Cela tient peut-être à son anglais, pour le moins rebutant. Mais je crois que la meilleure chose, c’est de rapporter fidèlement ce qu’il écrit lui-même à ce propos, en parlant — par exemple — de l’origine évolutive de la plume des Oiseaux.
« Comment se forma la plume ? De manière semblable à l’angiospermie, voilà ma réponse. Irais-je jusqu’à suggérer une cause ? Oui : ce fut un ‘choc métabolique’ qui agit, dans les deux cas, sur un substrat génétique manifestement chargé d’hérédité à fort rayonnement et très riche. Chez les Angiospermes, il semble que la ‘nouvelle’ embryogénie fasse référence à quelque chose de comparable aux algues : quelque chose qui n’est certainement pas là par effet de ‘descendance linéaire’. Suis-je alors un disciple de Goldschmidt qui croit à la ‘mutation systémique’ ? Les étiquettes ne m’intéressent pas, mais je suis sûr que nous devrons travailler beaucoup plus profondément et beaucoup plus intelligemment sur le problème fondamental de savoir ce qu’est le ‘gène’ et ce qu’est 1″hormone’ avant de pouvoir être tout à fait certains de nombreuses choses qu’il nous plaît de croire aujourd’hui évidentes (…) Et j’y insiste, je suis loin d’être certain que nous comprenions à présent ce qu’est le ‘gène’ et ce qu’est 1″hormone’, et encore moins de quelle façon ils interagissent » [29].
Concrètement, Croizat est donc obligé d’admettre qu’il ne sait pas comment fonctionne exactement le mécanisme de l’évolution. Il suppose que certaines expériences de type « mitchourinien » (du nom du lamarckien soviétique Mitchourine, prédécesseur de Lyssenko) — selon lesquelles des modifications profondes de la nutrition cellulaire entraîneraient parfois des variations héréditaires différentes des mutations classiques — peuvent avoir une certaine valeur, bien qu’elles aient été contestées. Ces variations héréditaires agiraient sur les contrôles génético-hormonaux d’une structure biologique préexistante et ayant un caractère « orthogénétique », pour reprendre l’adjectif employé par Croizat. L’écaille du poisson ou du reptile, la plume de l’oiseau et le poil du mammifère ne seraient en somme, pour Croizat, qu’autant de variations d’un seul thème fondamental « orthogénétique », c’est-à-dire structurellement capable de se modifier à la suite précisément de transformations génético-hormonales héréditaires induites par le milieu — dans le sens de ces variations (et, peut-être, d’autres possibles variations futures du même type), selon une direction déterminée. Ce qui revient à dire : dans un premier temps prédominait l’écaille, puis la plume et enfin le poil ; mais la réalité structurelle qui a permis la formation de ces différents appareils tégumentaires — réalité présente chez tous les Vertébrés — n’a fait que répondre différemment à plusieurs impulsions transformatrices, sans obéir à une finalité mystérieuse.
Je ne peux maintenant que vous poser la question quasi rituelle : que pensez-vous de toutes ces idées ?
En ce qui concerne les aspects biogéographiques de l’œuvre de Croizat, je les trouve fondamentaux. Ils nous enseignent à voir les espèces — comme l’avait déjà fait l’ornithologue allemand Otto Kleinschmidt [30] au début du siècle — non seulement comme des unités morphologiques, mais aussi comme des unités tout à la fois morphologiques, géographiques et chronologiques. Il en découle que la distribution géographique des espèces peut jouer un rôle essentiel dans leur identification exacte.
Je ne suis pas du tout convaincu, en revanche, par la façon dont Croizat conçoit le processus de transformation des vivants. Je crois que les facteurs géographiques et environnementaux peuvent certainement être déterminants dans la formation de races à l’intérieur d’une espèce et, probablement aussi, dans la différenciation d’une espèce en deux ou plusieurs espèces (en entendant par espèce ce qu’on entend communément : un ensemble de populations interfécondes) ; mais je ne crois pas que leur action puisse aller plus loin, jusqu’à ce qu’ils soient responsables, par exemple, de la différenciation d’un règne en types, d’un type en classes, d’une classe en ordres, d’un ordre en familles, d’une famille en genres. Je ne pense pas que la différenciation du règne animal en types aussi distincts que les Spongiaires, les Cœlentérés, les Mollusques, les Brachiopodes, les Annélides, les Arthropodes et les Échinodermes — différenciation qui existait déjà au début de l’ère paléozoïque — puisse s’expliquer par de simples facteurs de géographie sous-marine. Je ne crois pas à l’existence de populations d’organismes « plus généralisés » au sein desquelles se seraient différenciées, dans un deuxième temps et par vicariance géographique ou autrement, des populations « moins généralisées ». En d’autres termes, je ne crois pas au passage type -> classe -> ordre -> famille -> genre -> espèce, si on le conçoit comme un processus déterministe et linéaire s’étant vérifié sur le plan historique.
Cette analogie entre le processus de développement épigénétique individuel et l’histoire de la vie sur la Terre, autrefois proposée par Rosa et reprise aujourd’hui par Lovtrup et par presque tous les « cladistes », me semble totalement injustifiée et même aisément réfutable à la lumière des seules données qui permettent vraiment de trancher ici : celles que nous offre la documentation paléontologique.
La paléontologie nous montre sans équivoque que dans le passé les vivants constituaient un système organisé à plusieurs niveaux hiérarchiques, tout aussi grand et complexe que le système actuel.
Dans les mers du Cambrien, il n’existait pas d’Arthropodes pouvant être rapportés à un seul type morphologique « généralisé » au sein duquel, dans un deuxième temps, se seraient différenciées plusieurs classes qui, à leur tour, auraient donné naissance à plusieurs ordres, et ainsi de suite. En réalité, et tout comme aujourd’hui, les Arthropodes du Cambrien étaient « déjà » différenciés en classes, ordres, familles, genres et espèces. Ils formaient « déjà », autrement dit, un système hiérarchique bien identifiable à l’intérieur d’un système plus vaste, le règne animal.
Les divers plans hiérarchiques de complexité structurelle qui caractérisent chaque organisme vivant ne se sont pas réalisés en succession linéaire au cours du temps (du simple et de l’homogène au complexe et à l’hétérogène, comme l’imaginait Darwin, ou du général et hautement potentiel au particulier actualisé, comme le pensent Lovtrup et les « cladistes » orthodoxes) ; ils se présentent comme constamment superposés et même comme se compénétrant l’un l’autre à chaque moment de l’histoire de la vie. Seule une attitude relativiste permet donc de disposer en succession linéaire (ascendante ou descendante) tous ces plans de complexité structurelle qui, en réalité, se complètent harmonieusement et sont donc inséparables.
Relativement à notre point de vue particulier d’êtres humains, le Dobermans du voisin ou la chatte tigrée de notre ami seront, avant tout, Fido ou Fuffy. Mais ils seront en même temps (et vous voudrez bien excuser le ton pédant que je donne à mon propos, afin de me faire mieux comprendre) :
— d’une part, des systèmes d’organes, à leur tour systèmes de tissus, à leur tour systèmes de cellules, à leur tour systèmes d’organites, à leur tour systèmes macromoléculaires, à leur tour systèmes atomiques, à leur tour systèmes de particules, à leur tour systèmes de probabilités d’interaction quantique ;
— d’autre part, des systèmes appartenant à l’espèce Canis familiaris ou Felis catus, à leur tour systèmes appartenant aux genres Canis et Felis, à leur tour systèmes appartenant aux familles Canidae et Felidae, à leur tour systèmes appartenant à l’ordre Carnivora, à son tour système appartenant à la classe Mammalia, à son tour système appartenant au type Chordata, à son tour système appartenant au règne des Eucariota, …
… qui est à son tour partie intégrante de tout le Systema Naturae. Des systèmes partout, en somme !
Exactement. Aucune chose n’est séparable dans l’absolu, mais seulement par référence au système plus grand dont elle est partie intégrante.
Il y a, pour chaque cas, des différences considérables dans la recherche, selon qu’elle est dirigée dans un sens ou dans l’autre. Décomposer analytiquement les « holons » Fido et Fuffy dans leurs holons de niveau inférieur est relativement facile : il suffit de sectionner les deux animaux, d’isoler leurs organes, de découper ceux-ci en minces morceaux pour en distinguer les tissus respectifs, d’examiner au microscope électronique les cellules caractéristiques de chaque tissu, puis de décomposer aussi ces dernières dans leurs éléments organulaires, moléculaires et atomiques. Il sera par contre beaucoup plus difficile de remonter synthétiquement à la délimitation exacte des frontières qui définissent les plans structuraux de chien et de chat, de Canidé et de Félidé, de carnivore, de mammifère, de Cordé et d’eucaryote. Pour faire cela, en effet, une connaissance approfondie de tous les autres types d’organismes vivant sur notre planète est nécessaire : connaissance qu’on n’a pu acquérir qu’après des siècles d’intense travail de recherche scientifique, mais qui, aujourd’hui encore, n’est pas complète.
Analyse et synthèse, déduction et induction sont, on le sait, les deux pôles fondamentaux de l’activité gnoséologique. Mais elles ne peuvent pas être considérées comme des procédés totalement séparés et indépendants l’un de l’autre ; elles sont les directions opposées, mais complémentaires, d’un seul processus cognitif harmonieux. Aristote appelait épagogique ce processus par lequel, de façon circulaire, on descend de l’holos au meros pour remonter du meros à l’holos. Mais il est clair que tout cela suppose l’adoption d’un paradigme organiciste lié à une vision du monde comme réalité unitaire et « dynamiquement statique ».
Je crois qu’il ne manque pas grand-chose pour compléter l’analyse de la pensée évolutionniste contemporaine. Il ne nous reste à examiner, en effet, que le courant, fondé par des paléontologues anglo-saxons, et appelé école des punctuated equilibria, des équilibres ponctués. Les idées de ce courant sont désormais connues du grand public, grâce à l’infatigable activité de l’un de ses principaux fondateurs, Stephen Jay Gould de l’université de Harvard, auquel l’un des plus célèbres hebdomadaires internationaux, Newsweek, a consacré, le 29 mars 1982, sa page de couverture.
Pour l’école en question, les espèces seraient des unités biologiques très stables, donc caractérisées par une plasticité plutôt limitée. L’évolution biologique, par conséquent, n’adviendrait pas à travers un refaçonnement continu et progressif des espèces (comme l’a soutenu jusqu’à aujourd’hui, dans le sillage des idées de Darwin, la théorie synthétique), mais à travers des processus de « spéciation quantique » se déroulant, de manière totalement imprévisible, sur quelques dizaines de milliers d’années seulement, c’est-à-dire presque instantanément à l’échelle géologique. En somme, les espèces se formeraient de façon discontinue, en grand nombre et très rapidement, et tendraient ensuite à rester pratiquement inchangées pendant de longs millions d’années. Les lignes directrices évolutives les plus importantes (autrefois appelées aussi « orthogénétiques ») ne seraient que le résultat de soustractions ou d’additions d’espèces dues à la sélection agissant entre ces dernières (et non l’effet de la sélection agissant entre les individus composant les espèces elles-mêmes). Autrement dit, les espèces se formeraient sans arrêt dans le temps, mais seules celles possédant de plus grandes capacités de survie (dues au fait de disposer de structures ou caractères nouveaux et avantageux) et capables de produire plus rapidement des espèces postérieures, seraient préservées. Toutes les autres seraient par contre éliminées (figure 12).
J’aimerais savoir ce que vous pensez de ces thèses, qui relèvent de ce qu’on pourrait peut-être appeler un « néodarwinisme réformé ».
Si je peux parler avec franchise, je dirais que je suis stupéfait par la facilité avec laquelle se répandent aujourd’hui des idées qui, derrière des étiquettes elles-mêmes assez extravagantes, non seulement ne présentent aucune originalité (car elles n’ajoutent rien à ce qu’on savait déjà), mais se révèlent en outre, à la lumière d’une analyse approfondie, vraiment inconsistantes. C’est précisément le cas de la soi-disant « théorie » des équilibres ponctués ou intermittents, que vous avez très bien résumée.
Pourquoi dites-vous « soi-disant » théorie ? Ne mérite-t-elle pas ce nom à vos yeux ?
Elle ne le mérite pas du tout. Et si vous avez la patience de m’écouter encore un peu, je suis sûr que vous comprendrez aisément pourquoi. Les points les plus faibles de cette « théorie » sont aussi les plus importants et les plus décisifs, puisqu’ils concernent avant tout : 1) le processus de spéciation (avec l’apparition rapide de nouveaux caractères, propres à fournir les points de départ des lignes directrices évolutives postérieures) ; 2) le phénomène de la stase morphologique présentée par les espèces dans les longues périodes de temps qui suivent leur formation.
Comment a lieu la spéciation ? Dans un premier temps, l’idée que s’en faisaient Gould et les autres auteurs de la même école que lui, c’était, ni plus ni moins, celle de la spéciation allopatrique avancée par Mayr. Mais à présent, grâce aux ouvrages d’auteurs comme White, ils se sont rendus compte que la spéciation semble être un phénomène extrêmement complexe, explicable par plusieurs solutions, dont la solution allopatrique n’est qu’un cas particulier, ni certainement ni nécessairement le plus important. Je ne suis donc pas du tout surpris que les « ponctuationnistes », à commencer par Gould, aient fini par se déclarer de plus en plus agnostiques et peu intéressés par les modalités exactes qui auraient présidé à la formation des espèces. Actuellement, ils se disent prêts à reconnaître non seulement des mécanismes de spéciation allopatrique, mais aussi des phénomènes macromutationnels au niveau des chromosomes et des gènes régulateurs, et même des hopeful monsters du type envisagé par Goldschmidt.
31]. Dover fait remarquer que dans les organismes sexués, les gènes responsables de certains caractères, et beaucoup d’autres dont la fonction est inconnue, existent souvent en plusieurs couples (de deux à plusieurs milliers), constituant ainsi des familles multigéniques. Dans une espèce donnée x, la plupart des gènes d’une famille peuvent partager une certaine mutation, mais les même gènes de la même famille dans une autre espèce y, étroitement apparentée à x, ne peuvent pas partager cette même mutation. En d’autres termes, les membres de toutes les familles multigéniques (quelle que soit la dimension, fonction ou distribution de celles-ci dans les chromosomes) sont pratiquement homogènes et propres à chaque espèce. Pour Dover, ces unités homogènes discrètes au niveau génético-moléculaire qui caractérisent les espèces, ne sont pas le produit d’actions sélectives, mais viennent d’un flux continu d’échange et de renouvellement géniques qui se fait spontanément à travers le triple mécanisme de la conversion génique (deux gènes de la même famille, légèrement différents au début, finissent par devenir identiques), de l’échange inégal (deux chromosomes qui échangent des segments d’ADN de longueur inégale) et de la transposition (des segments d’ADN se reproduisent en plusieurs exemplaires et sautent sur un autre chromosome). Après une longue période de temps, lorsque la « poussée moléculaire » produite par le triple mécanisme aura fait que les gènes d’une famille, appartenant à un certain type A, auront tous été progressivement remplacés par des gènes d’un autre type B, l’espèce originelle se transformera en une autre espèce. Ainsi, les longues périodes de temps où agirait la « poussée moléculaire » et celles, très brèves, où, cette action étant terminée, de nouvelles espèces se formeraient, peuvent-elles correspondre aux longues et brèves périodes de stase morphologique et de spéciation des « ponctuationnistes ».
-
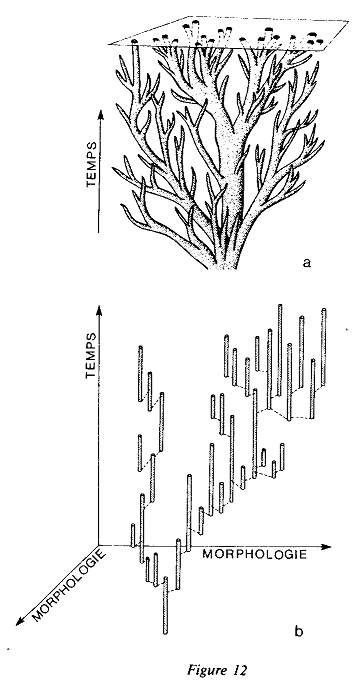
Le schéma de l’évolution biologique selon la conception darwinienne classique (en haut) et selon celle, plus récente, d’Eldredge et Gould, dite aussi des équilibres ponctués (en bas). Selon la conception classique, l’évolution des formes vivantes est un processus graduel qui se déploie comme les branches d’un immense arbre généalogique, chaque branche passant dans les branches successives sans solution de continuité et à travers chaque condition ou nuance intermédiaire. En conséquence, lorsqu’un paléontologue rassemble les fossiles d’un certain niveau stratigraphique, c’est comme s’il observait une « coupe » faite dans l’arbre de la vie (section transversale sur la figure en haut). À l’intérieur de cette « coupe », le paléontologue distingue plusieurs formes et les classe en différents taxa, en fonction du nombre de leurs ressemblances et/ou différences. On estime toutefois que ces taxa n’ont qu’une valeur conventionnelle, la seule réalité étant le flux unitaire et ininterrompu du processus d’évolution qui caractérise la vie.
Au contraire, selon le schéma des équilibres ponctués, l’évolution s’est déroulée, non tant de façon graduelle et uniforme qu’à travers de longues périodes de stase morphologique « ponctuées » de brèves périodes de transformation intense. En somme, les espèces (petits cylindres verticaux sur la figure en bas) resteraient fortement stables et essentiellement inchangées pendant des millions d’années — depuis leur apparition jusqu’à leur extinction dans l’échelle stratigraphique. Mais à certains moments de leur vie, sur des périodes de temps extrêmement brèves, et même « instantanées » du point de vue géologique (quelques milliers ou dizaines de milliers d’années), elles donneraient naissance à de nouvelles espèces, également stables et durables. Dans le modèle d’Eldredge et Gould, les espèces ne sont donc pas des conventions mais, en tant que quantités discrètes et identifiables, des choses réelles (schéma d’après Eldredge et Gould). Le modèle darwinien classique ne s’accorde pas avec les données paléontologiques ; mais celui des équilibres ponctués ne s’accorde avec celles-ci que pour ce qui concerne l’impossibilité de définir systématiquement les « chaînons intermédiaires » entre les taxa. Ce modèle, en effet, est tout à fait inadéquat et insuffisant pour rendre compte du phénomène grandiose des apparitions « explosives » et des extinctions brutales des faunes et des flores, phénomène qui s’est produit plusieurs fois au cours du temps géologique.
Il est clair que, pour Dover comme pour Kimura, la force motrice principale de l’évolution biologique n’est pas fournie par des facteurs déterministes et sélectifs, mais par des facteurs tout à fait spontanés et fortuits. Nous sommes donc en présence, une fois de plus, d’une conception très réductionniste, qui s’accorde mal avec les points de vue que j’ai développés et qui est, de toute façon, caractérisée par les mêmes insuffisances que celle d’Eldredge et de Gould.
Jusqu’à présent, la synthèse la plus élaborée et la plus détaillée des thèses soutenues par les « ponctuationnistes » est celle de Steven Stanley [32]. D’après cette synthèse, les individus constituant les espèces évolueraient incontestablement sous la poussée de la sélection naturelle, mais cette microévolution au niveau intradémique serait trop lente et trop faible pour réussir à produire des changements macroévolutifs, donc capables de mener à de nouvelles espèces. La macroévolution, elle, se produirait de manière imprévisible à la suite de l’émergence de nouveaux caractères, et en dépendance, aussi, des différents taux d’extinction connus par d’autres groupes de vivants. Admettre cela, en tout cas, revient à soutenir que la sélection naturelle agissant au niveau intraspécifique ne suffit pas à expliquer la macroévolution. L’extinction, en effet, ne rentre pas dans le domaine de la sélection naturelle (pour qu’il y ait sélection, rappelle Stanley, il faut qu’au moins quelques individus survivent et se reproduisent, tandis que dans le phénomène de l’extinction tous les individus d’une espèce cessent de survivre et de se reproduire), mais est provoquée par des facteurs limitatifs d’origine environnementale — comme la prédation, la compétition pour la nourriture et l’espace vital, l’influence du climat et de la quantité de nourriture à disposition, etc. — et par des facteurs fortuits. D’autre part, l’extinction d’une ou plusieurs espèces contemporaines ne peut pas ne pas jouer un rôle déterminant dans la survie et la reproduction de nouvelles espèces qui se forment pendant ou immédiatement après ladite extinction, car celle-ci met à la disposition des espèces nouvelles les niches écologiques laissées vides par la disparition, précisément, des premières espèces.
Or, le problème de fond est toujours le même. Si la sélection naturelle concernant les individus d’une espèce ne suffit pas à rendre compte de l’apparition de nouvelles espèces, et si la sélection entre espèces se limite à l’élimination ou à la préservation des espèces nouvellement constituées, comment se sont exactement formées ces dernières ? Placés face à ce problème, les « ponctuationnistes » manifestent alors toute leur impuissance, car : 1) ils se taisent ; 2) ou bien se déclarent, répétons-le, agnostiques et peu intéressés par la question ; 3) ou encore ils arrêtent la discussion en se contentant d’introduire, comme par exemple Stanley, le concept d’émergence de nouveaux caractères.
Si je ne me trompe, Konrad Lorenz recourt lui aussi à ce concept. À votre avis, a-t-il une certaine valeur ?
À mon avis, il n’en a aucune. Par le terme « émergence » on voudrait signifier qu’une qualité sort d’une chose sans que cette dernière la produise de la façon dont une cause produit un effet précis, la seule présence de la chose suffisant à expliquer l’apparition de cette qualité. Très peu de biologistes modernes, jusqu’à présent, ont recouru à cette notion; je ne me souviens en l’occurrence que de Conwy Lloyd Morgan, qui aspirait précisément, avec sa théorie de l’« évolution émergente » fondée essentiellement sur la philosophie de Samuel Alexander, à combiner l’évolution mécaniste de Darwin avec l’évolution vitaliste de Bergson. Selon Alexander, l’évolution produit de nouvelles structures et le seul fait de la présence de ces structures assure l’acquisition de nouvelles qualités auparavant inconnues. De ce point de vue, alors, la vie, le psychisme et Dieu ne sont que des propriétés de la matière (ou, comme dit Alexander, de l’espace-temps). On comprend donc aisément pourquoi d’autres auteurs — comme William McDougall [33] et Raymond Ruyer [34] — ont critiqué l’« émergentisme », y voyant une espèce de créationnisme à rebours, dilué et laïcisé.
Quoi qu’il en soit, il est évident que l’émergence n’est rien d’autre qu’un mot assigné à un phénomène admis a priori ; par là même, elle n’est pas l’explication de ce phénomène, pas plus que le mot « vie » n’explique la présence de la nutrition et de la reproduction. Ainsi que l’écrit André Lalande, « il est donc important de ne pas voir dans cette dénomination une hypothèse explicative, ni même une promesse d’intelligibilité » [35].
En effet. Du reste, elle ne suffirait pas non plus à rendre compte des phénomènes les plus grandioses de toute la paléontologie. Un paléontologue connu comme l’Anglais Chris Paul l’admet explicitement. Cette vision, écrit-il, « est inapte à expliquer l’apparition simultanée de nombreuses formes de vie (donc une radiation), ainsi que la disparition brusque de nombreuses espèces apparemment non reliées entre elles (une crise biologique) » [36].
Il y a peu, vous faisiez allusion au problème représenté par la « stase morphologique » traversée par les espèces au cours des longs millions d’années postérieurs à leur formation…
Cette question, elle aussi, n’est pas résolue par les « ponctuationnistes ». Deux partisans de la théorie synthétique, Ledyard Stebbins et Francisco Ayala [37], ont cru pouvoir l’aborder en introduisant la notion de « sélection stabilisatrice » ; mais il est clair que la vaste gamme des milieux occupés par des espèces actuelles ayant une morphologie uniforme en dépit d’une ample distribution géographique, ainsi que la stase morphologique à long terme (jusqu’à 17 millions d’années) manifestée par de nombreuses espèces fossiles qui vécurent dans des milieux instables et changeants, déposent fortement contre cette thèse. L’idée de Mayr [38] paraît plus convaincante : la stase morphologique de nombreuses espèces actuelles serait due à des limitations du développement imputables à une sorte d’équilibre homéostatique ; mais même dans ce cas, les mécanismes responsables de cet équilibre ne sont pas expliqués et restent donc obscurs, tout à fait hypothétiques. Il est évident, de toute manière, que si une forme quelconque d’homéostase devait se trouver à l’origine de la stase en question, alors le processus de spéciation devrait impliquer une destruction temporaire de cette homéostase au sein de l’espèce ancestrale. Et cela serait en accord avec ce que soutient Hampton Carson, que nous avons déjà rencontré à propos du problème de la spéciation.
Que dit exactement Carson ?
Il estime que les organismes sont gouvernés par deux systèmes génétiques superposés, dont l’un est « ouvert » et l’autre « fermé ». Le premier serait sensible à la sélection naturelle et donnerait lieu à des phénomènes d’adaptation intraspécifique à la suite de remplacements et de changements de fréquence des allèles. Le second, lui, extrêmement conservateur et insensible à la sélection naturelle, serait responsable des caractéristiques permanentes de chaque espèce en matière de développement, morphologie, physiologie et comportement. Carson nie que les processus de remplacement progressif des allèles puissent influencer le système fermé, et affirme que ce dernier ne peut mener à un nouveau système fermé spécifique — donc, concrètement, à une autre espèce — qu’à travers un phénomène « quantique » et discontinu, c’est-à-dire à la suite d’une série d’événements génétiques catastrophiques et casuels.
Récemment, le paléontologue Peter Williamson [39], étudiant les mollusques fossiles du Bassin du lac Turkana, en Afrique centre-orientale, a observé que les apparitions de nouvelles espèces étaient invariablement associées à des événements attestant une instabilité environnementale. Ces derniers ne pourraient-ils pas avoir contribué de façon décisive au démantèlement des « systèmes fermés » inhérents aux espèces ancestrales, menant ainsi à la formation des espèces nouvelles ?
J’ai du mal à l’admettre. Si les « systèmes fermés » sont vraiment insensibles à la sélection naturelle, comme l’affirme Carson, je ne vois pas de quelle façon les seules conditions extérieures pourraient arriver à les démanteler sans les détruire complètement. De toute façon, après avoir lu attentivement l’étude de Williamson, je dois dire que je ne suis pas convaincu que les données qui y sont recueillies démontrent de manière sûre la formation d’espèces nouvelles à partir d’espèces précédentes. Tout ce que ce travail a pu montrer, c’est que des sédiments plio-pléistocènes riches en mollusques du Bassin de Turkana indiquent un milieu lacustre qui est resté le même pendant des millions d’années. Ensuite, dans un intervalle compris entre 5000 et 50.000 années, le niveau du lac s’est brutalement abaissé et toutes les espèces de mollusques qui y avaient vécu pendant longtemps sans présenter aucun changement, subirent de grandes transformations dans la forme de la coquille. Williamson affirme que ces transformations étaient d’une importance telle qu’elles trahissaient la constitution d’une espèce nouvelle, mais les graphiques qu’il a joints à son étude ne suggèrent pas ce fait de manière absolument nette. Selon moi, les « espèces nouvelles » de Williamson pouvaient très bien être de simples normes de réaction aux conditions environnementales modifiées, normes qui se seraient manifestées dans les diverses espèces originelles. D’ailleurs, lorsque le niveau du lac remonta, les « espèces nouvelles » cessèrent d’exister, et les précédentes firent leur réapparition (avec quelques autres espèces très différentes, évidemment venues d’ailleurs). Williamson soutient que les données paléontologiques qu’il a étudiées — longues périodes de stase morphologique, « ponctuées » d’épisodes rapides de fort changement phénotypique — s’accordent avec le modèle d’Eldredge et de Gould. Mais ce modèle prévoit que les changements phénotypiques rapides mènent à la formation d’espèces nouvelles, qui devront ensuite rester longtemps identiques ; alors que dans le cas des mollusques du Turkana, il est évident que les « nouvelles espèces » (si elles étaient vraiment telles) s’éteignirent immédiatement après s’être formées.
Essayons maintenant de conclure au sujet de la conception des « équilibres ponctués ».
Somme toute, il me semble que les « ponctuationnistes » désirent avant tout mettre l’accent sur : 1) la stabilité des systèmes vivants et le caractère difficile de leur transformation ; 2) l’idée du changement évolutif comme transition rapide entre des états stables. À l’inverse, ils se détachent de la conception évolutionniste orthodoxe en niant : 1) qu’il y ait un « gradualisme phylétique » dans les phénomènes évolutifs (puisque les espèces fossiles se présentent comme bien individualisées et reconnaissables, dans la succession stratigraphique, dès leur première apparition, restant ensuite inchangées pendant longtemps) ; 2) que les possibilités inhérentes à la sélection naturelle s’exerçant à l’intérieur d’une population puissent être élargies au point de produire de nouvelles tendances évolutives et donc de nouvelles populations (ces dernières seraient, au contraire, le résultat d’une sélection entre des populations considérées comme des unités stables, c’est-à-dire entre des espèces proprement dites). En somme, la microévolution ne peut pas expliquer, à elle seule, la macroévolution.
En fonction de tout ce qui précède, on peut voir que la conception des équilibres ponctués n’a rien d’une véritable théorie scientifique : elle ne propose aucune explication originale et n’ajoute rien à ce qu’on savait déjà. Elle est plutôt le signe d’une attitude mentale nouvelle, qui marque la fin — chez les paléontologues — de l’état de sujétion où ils se trouvaient par rapport aux néontologues [40], longtemps réputés seuls susceptibles d’avoir voix au chapitre en ce qui concerne l’interprétation la plus appropriée des phénomènes évolutifs. Interprétation qui, évidemment, ne pouvait être, pour ces derniers, que la théorie synthétique.
Que les espèces soient des unités stables et qu’elles apparaissent à l’improviste dans la succession stratigraphique, on le savait déjà, en fait depuis la fondation de la paléontologie. Toutefois, à cause de l’évolutionnisme darwinien dominant, il était inconvenant, de mauvais goût et même dangereux (il y avait de quoi passer pour un réactionnaire obscurantiste !) de le dire publiquement et la tête haute. Comme le soutenait Darwin, l’absence des « chaînons intermédiaires » ne pouvait et ne devait être imputée qu’aux lacunes de la documentation fossile ! Mais l’interprétation darwinienne se révélant de plus en plus insuffisante, il était naturel qu’un tel état de choses cessât peu à peu. Chris Paul est un exemple éloquent de l’état d’esprit propre à la nouvelle génération de paléontologues, qui ne croient plus à l’inadéquation de la documentation fossile pour établir des généralisations théoriques sur l’origine de la vie.
Mais je voudrais que vous vous rendiez compte par vous-même des conclusions auxquelles arrivent les « ponctuationnistes », si l’on en juge par leurs plus récentes prises de position.
« Je ne veux pas soutenir la désespérante (sic) conclusion que la micro et la macroévolution sont absolument distinctes en règle générale et que rien de l’une ne peut éclairer l’autre », écrit Gould, qui ajoute : « Mais devons-nous accepter le ‘continuationnisme’ darwinien au risque d’ignorer les propriétés légitimement indépendantes de niveaux plus élevés que ceux où la sélection naturelle opère sur des organismes individuels ? Le choix doit-il impliquer désespoir et connaissance ou satisfaction et ignorance ? Je crois que la notion de hiérarchie résout ce faux problème, en intégrant les aspects positifs des deux positions. Les niveaux n’obtiennent pas leur indépendance du fait que quelque processus génétique fondamentalement nouveau apparaît. Ce sont plutôt les processus fondamentaux de variation et de sélection qui opèrent dans toute la hiérarchie. Mais ils travaillent de manière différente sur les divers matériaux des niveaux ascendants dans une hiérarchie discontinue. Nous pouvons apprendre tout ce qu’il nous faut savoir sur les lignes directrices évolutives en étudiant simplement ce qui se produit à l’intérieur des populations, pour la bonne et simple raison que les espèces peuvent agir comme unités de sélection. D’importantes relations de feed-back unissent tous les niveaux, mais de nouvelles modalités apparaissent à des niveaux plus élevés, et la réduction à la sélection naturelle agissant sur des organismes individuels ne pourra certainement pas rendre compte de tout dans l’évolution. L’équilibre ponctué (…) réclame la hiérarchie et remet en question le réductionnisme. C’est là son aspect le plus important dans la structure de la théorie évolutionniste » [41].
Ainsi donc, « on le voit », reconnaît Daniele Mazzonis dans sa préface à l’édition italienne du Pouce du panda, « le ‘démocrate’ et ‘marxiste’ Gould parle de hiérarchie et de holisme, et cherche ses arguments antiréductionnistes et tendanciellement ‘dialectiques’ dans le champ de l’hérésie anti-darwinienne et dans celui de l’organicisme biologique, traditionnellement réputés de droite » [42].
Les argumentations de Gould mettent fréquemment le cap vers les côtes de l’organicisme ; mais chaque fois que cela a lieu, le « holiste newyorkais » (c’est ainsi qu’il se définit), comme s’il était tout fâché, baisse pavillon et se laisse de nouveau porter par les tranquilles courants darwiniens, pour finir par s’en prendre à von Bertalanffy, à Koestler, à Capra (qu’il accuse de… réductionnisme !) et par continuer à proclamer à tous les vents la grandeur impérissable de Darwin, la fraîcheur et l’actualité de son œuvre. Comme Kimura, en effet, Gould reste un ardent darwinien, bien qu’il lui plaise de se définir aussi, plus précisément, comme un « néo-néodarwinien » (voulant ainsi faire entendre qu’il est allé plus loin que les partisans de la théorie synthétique, mais en respectant les tendances mutationniste-sélective ou casualiste-déterministe de cette dernière). Il submerge les lecteurs par un fleuve d’articles au ton cultivé et intelligent, brillant et désinvolte, souvent même frivole et moraliste, brouillant les cartes et laissant entendre que, même si la biologie devait un jour rejeter les formulations originelles de Darwin, le darwinisme ne mourrait pas pour autant, puisqu’il lui resterait la gloire éternelle de nous avoir convaincus de la réalité du « fait » de l’évolution [43]. Je suis souvent tenté de dire, à propos de Gould, que c’est un excellent biologiste de salon typiquement yankee ; mais puisqu’il s’agit aussi d’un collègue paléontologue, peut-être suis-je un peu trop méchant. [44]
Mais quels sont actuellement les biologistes qui, pour reprendre votre image, ont déjà mis le cap vers les côtes de l’organicisme ?
Je ne saurais faire ici un « recensement » suffisamment précis. Mais je peux vous assurer qu’ils sont nombreux et que leur nombre ne fait que croître. Il s’agit généralement de biologistes qui, à la différence de Gould, ont désormais rejeté définitivement la pensée darwinienne, ou bien qui en ont ouvertement dénoncé les lacunes, reconnaissant dans le processus évolutif un facteur « programmatique » ou « sémantique », c’est-à-dire — en dernière analyse — « intelligent » (bien qu’il soit ici question non d’une intelligence au sens anthropomorphique du terme, mais d’une intelligence propre à ce que j’appelais Esprit cosmique, Logica Entis ou Logica maior).
Rémy Chauvin, éthologiste, professeur à la Sorbonne et ancien élève de Grassé, soutient, par exemple, que la clé de l’évolution ne réside pas dans les « superstitions darwiniennes » mais, selon toute probabilité, dans un « programme » qui se réalise de la même façon que la manifestation de la volonté humaine, c’est-à-dire à travers des modalités dont les organismes n’ont pas conscience [45]. Il se rattache donc très clairement, lui aussi, à la « gnose de Princeton ».
La théorie « sémantique » de Marcello Barbieri, professeur de biologie moléculaire à l’université de Turin, est également très intéressante [46]. Puisqu’il s’agit d’un compatriote, et puisque des ouvrages comme le sien sont devenus, en Italie, aussi rares que les merles blancs, j’estime normal et juste d’en parler assez longuement.
Selon Barbieri, la vie ne peut pas être réduite — contrairement à ce qu’on affirme communément — à une dualité formée par le génotype (ADN, ensemble des instructions pour la construction des caractères de l’organisme) et le phénotype (protéines, ensemble des caractères qui sont l’expression des instructions génétiques), mais s’explique par une trinité formée du génotype, du ribotype et du phénotype (dans l’acception de Barbieri : le système formé par toutes les molécules fondamentales qui incluent le sucre ribosomique et, avant tout, les acides ribonucléiques : ARN messager, ARN de transfert et ARN ribosomique) qui semble jouer, à l’égard de la cellule, le même rôle essentiel que le cerveau par rapport au langage. En effet, de même que le cerveau aurait pour fonction sémantique de donner une signification aux informations recueillies par les sens et de les traduire en langage, de même le ribotype aurait pour fonction de donner une signification aux informations contenues dans l’ADN et de les traduire dans les expressions protéiques correspondantes. Ne pas tenir compte du ribotype, ce serait donc comme parler du langage en faisant abstraction du cerveau. Il s’ensuit, pour Barbieri, que l’origine de la vie doit être recherchée chez les précurseurs du ribotype, et que la dichotomie entre Procaryotes et Eucaryotes doit être ramenée à celle existant au niveau de leurs ribotypes. Sont ainsi mises à mal les autres hypothèses sur la genèse de la cellule vivante, à commencer par l’hypothèse orthodoxe selon laquelle les Procaryotes produisirent les Eucaryotes, qui à leur tour se réunirent en colonies pour donner naissance aux différents types d’organismes pluricellulaires.
En ce qui concerne l’origine brusque de la différenciation pluricellulaire au début de l’ère paléozoïque, Barbieri admet avec franchise que « c’est un problème pour lequel nous n’avons aucune solution (…) Sur le plan de la reconstruction historique, en conclusion, la théorie ribotypique des origines ne dépasse pas le Précambrien parce que la différenciation présente pour l’instant un obstacle insurmontable ». D’autre part, « sur le plan biologique, la théorie ribotypique de la cellule nous a déjà fourni les prémisses qui rendent concevable un mécanisme plus général de l’évolution » [47]. C’est ici précisément qu’est introduite une théorie « sémantique » de l’évolution.
Barbieri, tout d’abord, souscrit pleinement à la critique, faite par Lewontin, de la notion courante d’adaptation [48], reconnaissant qu’il y a interdépendance nécessaire entre organismes et niches écologiques, et que la formule darwinienne de la « survie du plus adapté » n’explique rien puisqu’elle n’est qu’une tautologie.
« L’idée que les organismes courent sans cesse derrière le milieu dans une chasse sans fin n’explique pas des événements dramatiques comme l’occupation de la terre par la mer et de l’atmosphère par la terre. Pour ces phénomènes, la théorie de Darwin réclame non seulement des organismes intermédiaires qui n’ont peut-être jamais existé, mais aussi des niches écologiques qui, à coup sûr, n’ont jamais existé ». Par ailleurs, « outre l’argument paléontologique que nous avons vu, il y a une objection de nature théorique à cette interprétation. Organismes et niches écologiques sont tellement interdépendants qu’il n’est pas possible de donner une définition réaliste de la niche sans les organismes qui l’occupent (…) Nous arrivons ainsi à une conclusion paradoxale. Si les niches ne peuvent être définies qu’en fonction des organismes qui les occupent, nous aurons des organismes et des niches parfaitement adaptés en vertu de leurs définitions mêmes, et dans ce cas il n’y a plus de place pour les processus d’adaptation. Nous dirons donc en conclusion que l’adaptation doit être parfaite et imparfaite en même temps : parfaite dans nos définitions et imparfaite dans la réalité. Un paradoxe, par conséquent, qui met en relief la nature intrinsèquement contradictoire de la notion darwinienne d’adaptation » [49].
Les choses étant ainsi, Barbieri se demande s’il n’est pas plus juste, alors, de dire que les organismes s’adaptent aux cycles naturels, plutôt qu’à leurs niches écologiques. En effet, « quand nous décrivons le cycle de l’oxygène, par exemple, l’atmosphère est traitée comme un membre du cycle, exactement comme les organismes. Être membre d’un cycle signifie simplement recevoir quelque chose d’un côté et donner quelque chose d’un autre côté, et sous cet angle organismes et niches écologiques se comportent de la même façon. Au lieu de dire que les organismes s’adaptent aux niches ou à l’environnement, nous pourrions donc dire que les organismes comme les niches s’adaptent ensemble aux cycles naturels, donnant ainsi naissance à des systèmes potentiellement capables de se perpétuer indéfiniment. De la sorte, nous nous apercevons qu’une alternative concrète à la notion darwinienne d’adaptation est en train de prendre forme : au lieu d »adaptation au milieu‘, nous commençons à parler d’‘adaptation aux cycles naturels » [50]. Imaginons maintenant, ajoute Barbieri, « un cycle naturel en voie de formation où il ne reste qu’une seule place pas encore occupée. Une espèce qui serait en mesure d’occuper cette place compléterait le cycle et apporterait une contribution importante, même si elle n’était pas bien adaptée à sa niche écologique. Autrement dit, avec le nouveau concept, le rôle d’une espèce dans l’économie de la nature est définie par sa participation aux cycles naturels, et non par son degré d’adaptation à sa niche écologique. Par là même, le concept d’adaptation parfaite n’est plus nécessaire, et le paradoxe, simplement, disparaît » [51].
Barbieri en arrive donc à la conclusion que l’évolution biologique n’a pas eu lieu seulement par sélection naturelle, mais aussi, et même surtout, en vertu de conventions naturelles : conventions qui trahissent certainement « un dessein, un grand principe de la nature » [52]. « Au concept d’adaptation au milieu, écrit-il, nous ajoutons le concept d’adaptation aux cycles naturels, et nous nous rendons compte que les organismes s’adaptent aux niches écologiques par sélection naturelle et aux cycles par conventions naturelles. Avec deux concepts d’adaptation, nous avons immédiatement deux mécanismes distincts de modification. On remarquera en outre que le nouveau mécanisme ne concerne pas des phénomènes mystérieux ou ambigus : les cycles naturels sont en permanence autour de nous, et nous sommes tous conscients de leur importance. Nous sommes donc en présence d’une nouvelle théorie de la vie : l’évolution biologique se produisit en vertu de conventions naturelles et de la sélection naturelle, de même que l’évolution linguistique eut lieu en vertu de conventions linguistiques et par sélection linguistique. C’est pour cette raison que je l’ai appelée théorie sémantique de l’évolution (…) (Elle) dépend de la théorie ribotypique de la cellule parce qu’elle a besoin de représenter la cellule comme une trinité de catégories fondamentales. Mais elle dépend aussi de la théorie ribotypique des origines parce que l’idée d’adaptation aux cycles naturels exige la reconstruction du cycle qui apparut en premier et qui fut la base de tous les autres cycles de la vie : le cycle cellulaire » [53].
À vrai dire, le parallèle établi entre l’évolution biologique et le langage humain n’est pas totalement original, puisqu’on peut déjà le trouver, exposé avec une certaine force, dans un livre du journaliste scientifique britannique Jeremy Campbell, paru il y a quelques années [54]. Mais ce qu’il est plus important de souligner, c’est que la théorie sémantique finit par renvoyer, du moins dans une large mesure, à la « grande chaîne de l’Être » d’Aristote et au Systema Naturae de Linné et de Cuvier. Barbieri écrit en effet que « les cycles sont stables, potentiellement immortels et se disposent en hiérarchies de cycles à l’intérieur d’autres cycles ; surtout, ils nous disent que tous les organismes sont complémentaires, un concept aussi vieux que la civilisation. Les biologistes en ont toujours été conscients, mais dans le système de référence darwinien, où compétition et sélection sont les lois suprêmes, l’idée de complémentarité devient à peine croyable. Ceci a été la pierre de touche qui a divisé les scientifiques (…) Sur ce point, la théorie sémantique s’oppose au darwinisme et à la synthèse moderne. La loi suprême de la vie, c’est la coopération et l’interdépendance de toutes les créatures, non la compétition et la sélection. La compétition est incontestablement une donnée de fait, mais l’adaptation aux cycles est fondamentale. Avant toute chose et au-dessus de toute chose, les formes de la vie doivent créer des cycles naturels coopérant entre eux, se répartissant les tâches et remplissant des fonctions complémentaires. Après quoi chaque place dans les cycles peut être occupée par des créatures légèrement différentes, et une certaine compétition entre elles pour s’en assurer la possession est naturelle. Mais il s’agit d’événements secondaires au sens littéral du terme : sans les événements primaires qui ont mené à la formation des cycles, les processus de compétition et de sélection ne pourraient même pas commencer à exister » [55].
Puisque les cycles naturels, tels que nous les décrit Barbieri, sont de véritables systèmes cybernétiques, où chaque élément interagit de manière étroite avec tous les autres, je trouve cette théorie « sémantique » beaucoup plus en accord avec le paradigme organiciste qu’avec le paradigme évolutionniste.
Il me semble qu’elle ouvre aussi une piste importante pouvant mener à une meilleure compréhension des systèmes écologiques en général.
Sans doute. À ce sujet, il est même nécessaire de rappeler que la vision darwinienne de la nature comme « crocs et griffes rouges de sang », jungle impitoyable où compétition et prédation sont les deux thèmes dominants, a été sérieusement remise en cause au cours des dix dernières années. À la place de cette vision s’est répandue de plus en plus l’idée que le mutualisme — ensemble d’interactions mutuellement bénéfiques entre les espèces vivantes — joue un rôle aussi important que la compétition et la prédation [56].
Mais c’est surtout dans deux livres importants parus presqu’en même temps que celui de Barbieri, que l’organicisme est vraiment mis en relief. Le premier est un recueil d’articles de plusieurs auteurs critiques à l’égard de la théorie synthétique, recueil publié sous la direction de Mae-Wan Ho, de la Open University Milton Keynes, et de Peter T. Saunders, de l’université de Londres, sous le titre significatif de Beyond Neo-Darwinism. An Introduction to the New Evolutionnary Paradigm. [57] Pour savoir en quoi consiste également ce « nouveau paradigme », il suffit de parcourir la préface de l’ouvrage, où l’on peut lire les phrases suivantes : « Jusqu’à il y a quelques années seulement, la théorie ‘synthétique’ ou ‘néodarwinienne’ de l’évolution se présentait, virtuellement inattaquable, comme le fondement de notre compréhension du monde organique. Certains, à vrai dire, restaient en dehors du consensus général, mais exerçaient très peu d’influence sur la majorité des biologistes. Presque toute la recherche qui était faite sur l’évolution était programmée dans le but d’étudier les modalités d’action de la sélection naturelle, et était censée confirmer obligatoirement la théorie. Mais aujourd’hui le paysage est complètement différent. Un nombre toujours croissant d’auteurs se déclarent insatisfaits par la théorie synthétique (…) Tous ces signes dénotent que la théorie est en crise et qu’un changement se prépare. En outre, on peut déjà prévoir le type de changement qui s’annonce (…) La chose la plus importante à souligner au sujet de l’alternative qui se présente, c’est qu’il ne s’agit pas d’une nouvelle théorie, comme voudrait le faire croire le néodarwinisme, mais d’un paradigme. C’est une façon d’aborder les problèmes, plutôt qu’une solution. La théorie synthétique est fondée sur une explication définitive de la façon dont se produit l’évolution. On suppose que l’évolutionniste l’accepte comme une donnée de fait, se contentant simplement de montrer comment le processus peut s’être déroulé dans des cas différents entre eux. La théorie a aussi une grande influence sur la manière dont les organismes sont étudiés. L’effet le plus important, c’est la tendance à considérer les organismes et même les sociétés comme des entités décomposables en morceaux distincts, dont chacun doit nécessairement conférer quelque avantage sélectif. Par contre, dans le cadre du nouveau paradigme, on commence par étudier les organismes et les sociétés comme s’ils étaient des totalités intégrées. Après quoi l’on cherche à utiliser ce qu’on a appris par cette étude pour arriver à mieux comprendre comment ils ont évolué » [58].
Une approche tout à fait identique inspire le deuxième ouvrage, écrit par le biologiste canadien Robert G.B. Reid, de l’université de Victoria, dont l’un des principaux objectifs est de montrer que le schisme entre la vision quasi mystique de l’évolution et l’étroite vision positiviste ne sert qu’à préparer le terrain aux théories pseudo-scientifiques les plus disparates, alors qu’une approche « plus holiste » peut mener à une synthèse de philosophies opposées entre elles [59]. Chose très significative : deux chapitres du livre s’intitulent respectivement « Holisme et biologie » et « Holisme et évolution », comme pour confirmer que l’ouvrage se place bien dans le sillage des travaux de J.C. Smuts et de bon nombre d’autres biologistes européens d’avant la deuxième Guerre mondiale.
___________________________________________________________________
1 Cité dans l’ouvrage collectif Mutamenti e continuità nelle scienze umane e naturali, «Elite», Firenze, 1982, p. 38.
2 Cf. G. de Beer et W.E. Swinton, Prophetic Fossils, in T.S. Westoll (ed.), Studies on Fossil Vertebrates, Athlone Press University, London, 1958 ; G. de Beer, Embryos and Ancestors, Oxford University Press, Oxford, 1958 ; Gareth Nelson, Ontogeny, Phylogeny, Paleontology and the Biogenetic Law, in Systematic Zoology, XXVII, 3, 1978, pp. 324-345.
3 Cf. B. Kurten, Inte fran aporna, Bonniers Forlag AB, Stocklom, 1971 (trad. it. : Non dalle scimmie, Einaudi, Torino, 1972).
4 Stephen Jay Gould, Ontogeny and Phylogeny, Harvard University Press, Cambridge (Mass.), 1977.
5 Soren Lovtrup, On von Baerian and Haeckelian Recapitulation, in Systematic Zoology, XXVII, 3, 1978, pp. 348-352.
6 Soren Lovtrup, The Four Theories of Evolution, in Rivista di Biologia, 75, 1-2-3, 1982.
7Soren Lovtrup, Art. cité, p. 61.
8 Richard Goldschmidt, The Material Basis of Evolution, Yale University Press, New Haven, 1940.
9 Otto H. Schindewolf, Grundlagen der Paläontologie, Schweizerbart, Stuttgart, 1950.
10 Albert Dalcq, Initiation à l’embryologie générale, Desoer, Liège, 1952.
11 Soren Lovtrup, Art. cité, p. 260.
12 Giuseppe Sermonti, On Lovtrup’s Approach to Evolution, in Rivista di Biologia, 75, 4, 1982.
13 Richard C. Lewontin, The Genetic Basis of the Evolutionary Change, Columbia University Press, New York, 1974, p. 159.
14 « Au milieu du chemin de notre vie / je me trouvai par une selve obscure / et vis perdue la droiturière voie. / Ha, comme à la décrire est dure chose / cette forêt sauvage et âpre et forte, / qui, en pensant, renouvelle ma peur ! » (Dante, Œuvres complètes, trad. André Pézard, La Pléiade) (N.D.T.).
15 On appelle hétérozygote une cellule ou un organisme diploïde (c’est-à-dire dans lesquels chaque chromosome est présent en deux exemplaires) qui possède deux allèles différents d’un ou de plusieurs gènes, et homozygote une cellule ou un organisme diploïde qui possède deux allèles identiques du ou des gènes considérés.
16 Motoo Kimura, The Neutral Theory of Molecular Evolution, Cambridge University Press, Cambridge, 1983. Pour un exposé plus succinct de cette théorie, voir l’article (même titre que l’ouvrage) publié par Kimura dans The New Scientist du 11 juillet 1985.
17 J. Gribbin et J. Cherfas, The Monkey Puzzle, 1982.
18 R.C. Richmond, Non-Darwinian Evolution : a Critique, in Nature, 225, 1970, pp. 1025-1028.
19 Masatoshi Nei, Molecular Population Genetics and Evolution, North-Holland Publ. Co., Amsterdam-Oxford, 1975, p. 249.
20 Georges Pasteur, Génétique biochimique et populations, ou : pourquoi sommes-nous multipolymorphes? in Le polymorphisme dans le règne animal, « Mémoires de la Société Zoologique de France », 36, 1972, p. 501.
21 Alister Hardy, The Living Stream, Collins, London, 1965.
22 Hampton L. Carson, The Genetics of Speciation at the Diploid Level, in American Naturalist, 109, 1975, pp. 83-92.
23 Cf. par exemple : V.C. Wynne-Edwards, Animal Dispersion in Relation to Social Behavior, Oliver and Boyd, London, 1962. Cet auteur a démontré, scandalisant les darwiniens orthodoxes, que de nombreuses espèces animales exercent un contrôle sur leurs possibilités reproductives et maintiennent leurs populations à des niveaux constants, comme en accord avec les ressources de nourriture disponibles.
24 Ernst Mayr, Animal Species and Evolution, Harvard University Press, Cambridge (Mass.), 1963.
25 Cf. Guy L. Bush, Modes of Animal Speciation, in Annual Review of Ecology and Systematics, 6, 1975, pp. 339-369 ; et What Do We Really Know about Speciation ? in R. Milkman (ed.), Perspectives on Evolution, Sinauer, Sunderland, 1982, pp. 119-128.
26 Michael J.D. White, Modes of Speciation, Freeman, San Francisco, 1978.
27 Léon Croizat, Space, Time, Form. The Biological Synthesis, édité par l’auteur, Caracas, 1962.
28 Cf. Gareth Nelson et Donn Eric Rosen, Vicariance Biogeography, Columbia University Press, New York, 1981.
29 Léon Croizat, Op. cit., pp. 452-453.
30 Otto Kleinschmidt, Die Formenkreislehre und das Weltwerden des Lebens, Gebauer-Schwetscke, Halle, 1926 (trad. anglaise: The Formenkreis Theory and the Progress of the Organic World, Witherby, London, 1930).
31 Gabriel Douer, Molecular Drive : a Cohesive Mode of Species Evolution, in Nature, 299, 1982, pp. 111-116 ; et, du même auteur, Le forte dell’evoluzione, in « Kos », 5, 1984, pp. 129-136.
32 Niles Eldredge et Stephen Jay Gould, Punctuated Equilibrium. An Alternative to Phyletic Gradualism, in T. Schopf (ed.), Models in Paleobiology, Freeman, San Francisco, 1972, pp. 82-115 ; voir aussi : Steven M. Stanley, Macroevolution. Pattern and Process, Freeman, San Francisco, 1979.
33 William McDougall, Modern Materialism and Emergent Evolution, Methuen, London, 1934.
34 Raymond Ruyer, Néo finalisme, PUF, 1952.
35 André Lalande, Vocabulaire technique et critique de la philosophie, PUF, 1980, p. 277.
La vision « ponctuationniste » se révèle donc plutôt inconsistante.
36 Chris Paul, The Natural History of Fossils, London, 1980.
37 Ledyard Stebbins et Francisco Ayala, Is a New Evolutionary Synthesis Necessary ? in Science, 213, 1981, pp. 967-971.
38 Ernst Mayr, Population, espèces et évolution, Hermann, 1974.
39 Peter G. Williamson Paleontological Documentation of Speciation in Cenozoic Molluscs from Turkana Basin, in Nature, 293, 1981, pp. 437-443.
40 Pour certains biologistes, la paléontologie, en tant que science fondée sur l’étude des fossiles, ne peut pas être considérée comme une branche de la biologie. Pour Roberto Fondi et d’autres auteurs, elle l’est de plein droit ; il faut donc entendre ici par « néontologues » tous ceux qui étudient les espèces vivantes actuelles (N.D.T.).
41 Stephen Jay Gould, Punctuated Equilibrium. A Different Way of Seeing, in The New Scientist du 15 avril 1982 (souligné par nous).
42 Stephen Jay Gould, Le pouce du panda. Les grandes énigmes de l’évolution, Grasset, 1982 (références de la trad. it. : Il pollice del panda, Editori Riuniti, Roma, 1983).
43 Gould se montre souvent irrité par les « créationnistes scientifiques » de son pays, parce qu’ils se permettent de présenter ses thèses comme des exemples éloquents d’antidarwinisme. Mais sous l’effet de la colère, il se laisse parfois aller à des affirmations qui ne manquent pas d’être étonnantes. Voici deux exemples : « Les formes transitoires manquent habituellement au niveau des espèces, mais elles abondent entre groupes plus vastes » (Quand les poules auront des dents. Réflexions sur l’Histoire naturelle, Fayard, 1984, p. 285). Tous les paléontologues, à commencer par l’ultra-darwinien Simpson, savent que c’est exactement le contraire qui est vrai. « Les faits sont les données de l’univers. Les théories sont des structures d’idées qui expliquent et interprètent les faits. Les faits ne disparaissent pas quand les scientifiques débattent de théories antagonistes qui prétendent les expliquer. La théorie de la gravitation d’Einstein a remplacé celle de Newton, mais les pommes ne s’immobilisent pas au beau milieu de leur chute en attendant que le débat soit tranché. Et les êtres humains ont évolué à partir d’ancêtres qui ressemblaient à des singes, qu’ils l’aient fait en fonction de l’explication proposée par Darwin ou d’un autre mécanisme qui reste encore à découvrir » (op. cit., p. 279). Un créationniste pourrait facilement répondre à Gould que tout le monde, ou peu s’en faut, a déjà vu des pommes tomber des arbres, mais que personne n’a jamais vu un singe se transformer en homme. Le paléontologue de Harvard confond donc, précisément, les faits (ou phénomènes) avec les théories.
44 Il est péremptoirement affirmé, sur la quatrième page de couverture de l’édition française du livre Le pouce du panda, que Stephen Jay Gould est « unanimement considéré comme le plus brillant paléontologue de notre époque »… (N.D.T.).
45 Rémy Chauvin, La biologie de l’esprit, Rocher, Monaco, 1985.
46 Marcello Barbieri, La teoria semantica dell’evoluzione, Boringhieri, Torino, 1985. Trad. anglaise : The Semantic Theory of Evolution, Harwood Academic Publishers, London-New York, 1985 (avec une préface de René Thom).
47 Marcello Barbieri, Op. cit., p. 167.
48 Lewontin, professeur de génétique à l’université de Harvard, a fait remarquer que « les organismes n’expérimentent pas les milieux passivement, mais créent et définissent le milieu où ils vivent (…) Il y a une interaction constante entre organisme et milieu, de sorte que même si la sélection naturelle peut faire s’adapter l’organisme à une série particulière de circonstances environnementales, l’évolution de l’organisme modifie à son tour ces circonstances. Les organismes eux-mêmes déterminent, enfin, par leur activité, quels facteurs externes devront faire partie de leur niche ». Mais alors, « si les niches écologiques ne peuvent être spécifiées que par les organismes qui les occupent, l’évolution ne peut plus être décrite comme un processus d’adaptation, parce que tous les organismes sont déjà adaptés ». Par ailleurs, « si la sélection naturelle améliorait vraiment la capacité d’adaptation des organismes à leurs milieux, alors on pourrait s’attendre à ce que la probabilité d’extinction dans un proche avenir soit moindre pour les espèces qui existent déjà depuis longtemps, étant donné qu’on peut penser qu’elles ont été améliorées par la sélection naturelle. Toutefois, les données montrent que la probabilité d’extinction d’une espèce est apparemment une constante caractéristique du groupe auquel elle appartient, mais indépendante du temps, long ou bref, de son existence sur la Terre. Autrement dit, il ne semble pas que la sélection naturelle augmente, à long terme, les probabilités de survie d’une espèce ; elle lui permet simplement de se laisser remorquer ou de marcher au même pas que le milieu qui change constamment » (Richard C. Lewontin, L’adattamento, in Pietro Omodeo (ed.), Storia naturale ed evoluzione, Le Scienze, Milano, 1979, pp. 41-42 ; souligné par nous). Malgré ces observations tout à fait hétérodoxes, Lewontin ne songe même pas à abandonner la philosophie darwinienne de la sélection des mieux adaptés, car cela reviendrait à « jeter l’enfant avec l’eau du bain ».
49 Marcello Barbieri, Op. cit., p. 173.
50 Op. cit., p. 176.
51 Op. cit., p. 178. Souligné par nous.
52 Op. cit., p. 176. Souligné par nous.
53 Op. cit., p. 188 et p. 189.
54 Jeremy Campbell, Grammatical Man. Information, Entropy, Language and Life, Simon and Schuster, New York, 1982.
55 Marcello Barbieri, La teoria. semantica dell’evoluzione, op. cit., p. 192. Souligné par nous.
56 Cf. à ce sujet : D.H. Boucher (ed.), The Biology of Mutualism : Ecology and Evolution, Academic Press, London, 1985.
57 Mae-Wan Ho et Peter T. Saunders (ed.), Beyond Neo-Darwinism. An Introduction to the New Evolutionary Paradigm, Academic Press, London, 1984.
58 Mae-Wan Ho et Peter T. Saunders, Op. cit., pp. IX-X. Souligné par nous.
59 Robert G.B. Reid, Evolutionary Theory : the Unfinished Synthesis, Academic Press, London, 1984.