
Nous sommes donc arrivés à Dopo Darwin [1], le livre écrit par le professeur Sermonti [2] et vous-même. Paru en 1980, cet ouvrage qui en est déjà à sa quatrième édition a fait beaucoup de bruit et vous a valu des critiques féroces dans le monde universitaire. En premier lieu, estimez-vous que les deux positions opposées, évolutionniste et antiévolutionniste, sont le fruit de choix prélogiques, déterminés par la conception du monde de chacun, et qu’on cherche ensuite à rationaliser scientifiquement, ou bien sont-elles plus simplement le produit de cultures et de formes mentales différentes ?
Je dirais que les deux choses ne s’excluent pas, car il est naturel qu’à des visions du monde différentes correspondent des formes mentales et des cultures différentes. Il est tout aussi naturel que lorsque prédomine une certaine vision du monde et que, par sa faute, des signes d’insatisfaction se manifestent, certains adoptent une attitude critique, et non plus d’acceptation passive. Cela, non seulement par simple esprit anticonformiste, mais aussi pour éprouver la validité effective de cette vision et, dans le même temps, pour s’interroger sur leurs propres insatisfactions. Quoi qu’il en soit, nous ne devons pas oublier que la science a toujours un caractère duel. Elle est le résultat d’un dialogue ininterrompu entre l’homme et la nature. Et puisque l’homme, je le répète, n’est pas le simple objectif d’un appareil photographique qui transfère mécaniquement sur la pellicule l’image de ce qui se tient devant lui, mais un être pensant qui a d’innombrables problèmes — théoriques et pratiques — à affronter et à résoudre, alors il est logique et inévitable qu’il doive interroger intentionnellement la nature, c’est-à-dire en fonction de ses problèmes. Il est clair que si l’homme a des intentions bien définies et n’est pas disposé à tricher, le dialogue avec la nature sera pour lui profitable et constructif. Dans le cas contraire, nous aurons des réponses ambiguës et indifférentes. Eh bien, les intentions « traditionnelles » — qui visaient à découvrir dans la réalité une totalité organique et non un chaos de singularités indépendantes les unes des autres — n’ont pas été déçues par le dialogue avec la nature. En revanche, les intentions évolutionnistes — qui visaient à reconduire chaque chose au déterminisme et au hasard — ont été systématiquement démenties.
Pourriez-vous résumer ici, dans ses grandes lignes, le contenu de votre critique de l’évolutionnisme, telle qu’elle a été développée dans Dopo Darwin ?
Bien sûr. Comme vous le savez, selon l’idée évolutionniste courante n’importe quel organisme vivant — du grillon au faisan, et du pavot à l’homme — ne serait autre que la résultante d’un très long processus de transformation progressive, entamé il y a des centaines de millions d’années et menant de formes unicellulaires à des formes pluricellulaires de plus en plus complexes, jusqu’à culminer dans les mammifères et dans l’homme. Comme si cela ne suffisait pas, l’idée est appliquée à l’origine même des êtres vivants, qui se seraient formés à partir de la matière brute, suite à un long jeu de combinaisons chimiques de type classique entre des molécules organiques dispersées dans les eaux primordiales de notre planète.
Les preuves les plus évidentes de la validité, ou de l’invalidité, de l’idée évolutionniste doivent venir — non uniquement mais surtout — de la science des fossiles, la paléontologie, et de la géologie stratigraphique, qui se fonde sur la première. Comme le reconnaissait aussi le grand zoologiste Pierre-Paul Grassé, la paléontologie, en effet, est « la seule vraie science de l’évolution » [3]. Il est clair en effet que si le processus en question s’était réellement vérifié, l’histoire de notre planète 1) devrait nous présenter une très longue période initiale privée de vie et correspondant la phase d’« évolution chimique « dans les eaux primordiales ; 2) devrait nous faire voir des successions de roches sédimentaires de plus en plus riches en organismes unicellulaires fossilisés ; 3) devrait donc nous faire assister à la genèse des premiers organismes pluricellulaires rudimentaires (composés de quelques cellules seulement) ; et enfin 4) devrait nous montrer la naissance d’organismes pluricellulaires de plus en plus complexes et nombreux, jusqu’aux Arthropodes (araignées, insectes, etc.), aux Vertébrés et à l’homme. En outre, 5) les différentes formes vivantes devraient pouvoir se ranger sans difficulté dans un grand arbre généalogique, où chaque branche passe dans les suivantes sans solutions de continuité et à travers chaque situation intermédiaire. Mais tout cela, est-ce vraiment le tableau de l’histoire biologique que nous offrent la géologie stratigraphique et la paléontologie ? Malheureusement pour les évolutionnistes, si l’on admet que la succession stratigraphique construite par la géologie moderne reflète vraiment l’histoire de la vie sur notre planète, alors il faut répondre de manière négative.
Le fait est que déjà dans les roches sédimentaires les plus anciennes, comme par exemple celles d’Isua au Groenland et celles de Pilbara en Australie (dont l’ancienneté est estimée, respectivement, à 3,8 et à 3,5 milliards d’années, et qui appartiennent donc aux époques qui suivirent de près la consolidation et le refroidissement de la croûte terrestre, survenus, estime-t-on, il y a environ 4 milliards d’années), on trouve des corps microscopiques de substance organique à forme sphéroïdale ou à bâtonnet, de même que des structures sédimentaires lamineuses (dites stromatolites), lesquels sont généralement attribués à des fossiles de bactéries ou d’algues bleues, ou bien à l’activité biologique de ces dernières. Et c’est aussi un fait que des structures microscopiques sphéroïdales, à bâtonnet ou filamenteuses tout à fait analogues à celles découvertes dans de nombreuses roches pré-paléozoïques sont assez souvent présentes — comme l’a récemment démontré le paléontologue Hans Dieter Pflug, de l’université Justus Liebig de Giessen — dans des météorites carbonées tout aussi vieux.
Si l’attribution de toutes ces microstructures à des restes ou à des productions de véritables organismes unicellulaires est juste, nous devons en déduire que la vie est apparue à la surface de la Terre sous une forme « déjà prête », dès que les conditions environnementales de celle-ci furent aptes à l’accueillir. Peut-être la vie sera-t-elle venue en pluie des espaces extraterrestres bord de comètes ou de météorites (comme l’affirment les astronomes Fred Hoyle et Chandra Wickramasinghe [4], ou carrément dans des vaisseaux spatiaux étrangers (comme le soutient le généticien Francis Crick [5]) ? Personne ne peut le savoir. De toute façon, il n’en reste pas moins que la longue période sans vie supposée par les évolutionnistes et nécessaire à l’accomplissement de l’« évolution chimique » , fait totalement défaut.
Et si les microstructures auxquelles nous avons fait allusion n’étaient pas du tout des restes d’organismes vivants, mais de purs et simples lusus naturae, des « plaisanteries de la nature », ayant donc une origine purement minérale ? Même dans ce cas, les choses ne se présenteraient pas mieux pour les évolutionnistes. La documentation stratigraphique, en effet, montre que ces structures microscopiques ont survécu, pratiquement inchangées, dans les eaux des ères pré-paléozoïques, pendant environ 3 milliards et demi d’années ; après quoi s’est produit, à partir du début du paléozoïque (il y a environ 600 millions d’années), le phénomène le plus inquiétant de toute la paléontologie. Tout à coup (géologiquement parlant) sont apparues des quantités considérables de fossiles, restes pétrifiés de formes de vie pluricellulaire pouvant être parfaitement rapportées aux grands plans d’organisation connus de tout étudiant en zoologie : Éponges, Cœlentérés, Brachiopodes, Mollusques, Annélides, Arthropodes, Échinodermes, et d’autres que nous ignorons totalement parce qu’elles ont disparu en cours des ères suivantes.
Il est clair que, des énigmatiques corps microscopiques contenus dans les roches les plus anciennes et dans les météorites carbonées à ces restes incontestables d’organismes à la complexité anatomique souvent déconcertante (il suffit de penser aux fameux trilobites), le saut est considérable (figure 3). Et ce saut n’est pas explicable selon les schémas prévus par l’idée évolutionniste ; tant il est vrai qu’aucun partisan de celle-ci — à commencer par Darwin lui-même — n’est jamais parvenu à en donner une explication convaincante. D’innombrables hypothèses ont été avancées à ce sujet, mais aucune n’a pu résister à une critique approfondie.
|
Figure 3 Illustration du passage brutal entre la probable vie unicellulaire présente dans les formations rocheuses pré-paléozoïques (Bactéries, Cyanophycées et, dans un deuxième temps, Algues à fonction chlorophyllienne) et la vie pluricellulaire attestée, présentant une grande complexité, apparue au début de l’ère paléozoïque. Parmi les exemples d’organismes pluricellulaires infra-paléozoïques (fossiles des sédiments d’Ediacara, en Australie), figurent ici : un probable ver plat à symétrie bilatérale, Dickinsonia (en haut à gauche) ; un cœlentéré anthozoaire de l’ordre des Pennatules, Charnia (en haut au centre) ; un probable arthropode trilobitomorphe, Spriggina (en haut à droite) ; un autre probable arthropode de la classe des Crustacés, Parvancorina (en bas à gauche) ; un cœlentéré scyphozoaire, Cyclomedusa (en bas au centre) ; un probable échinoderme, Tribrachidium (en bas à droite). |
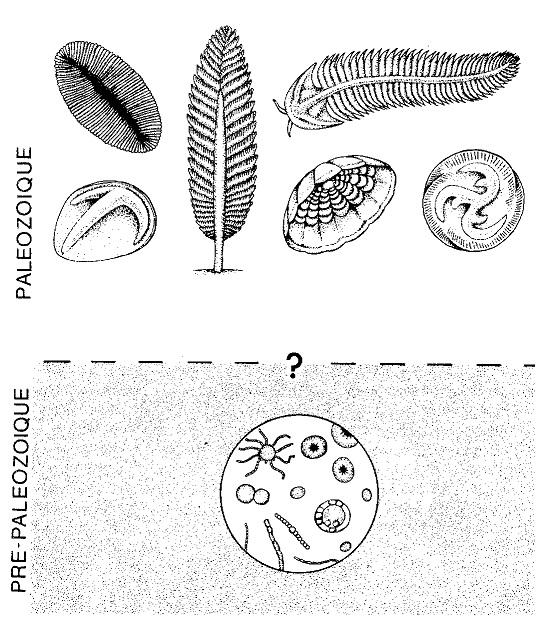
Mais ce n’est pas tout. Du début du paléozoïque à aujourd’hui, l’histoire de la vie n’apparaît pas comme comparable à celle d’un immense arbre généalogique, produisant — progressivement et sans interruption — des branches de plus en plus complexes et nombreuses. Elle est au contraire dominée par ce que bon nombre de paléontologues anglo-saxons, dont les chefs de file sont Niles Eldredge et Stephen Jay Gould, appellent des « équilibres ponctués » (punctuated equilibria), et que l’anthropologue Alfredo Sacchetti et moi-même nommons des seuils d’équilibre taxologique [6]. C’est une histoire faite de discontinuités structurelles (fût-ce au sens statistique ou probabiliste du terme) et d’équilibres homéostatiques entre ces discontinuités (qui trahissent des rapports interrelationnels à l’intérieur d’un véritable Systema biologique naturel) : une histoire où l’on assiste, périodiquement et sur une grande échelle, à des apparitions à l’improviste de formes nouvelles, souvent radicalement différentes entre elles, formes qui demeurent essentiellement inchangées pendant des millions et des millions d’années, avant de s’éteindre de façon tout aussi brutale et d’être immédiatement suivies de nouvelles apparitions.
Les fameux « chaînons manquants », ou « ancêtres communs » aux différentes catégories systématiques (taxa) des règnes végétal et animal, dont toute la documentation fossilifère devrait cependant être remplie — d’après l’idée évolutionniste — ne trouvent pratiquement aucune place dans ce tableau. Et en effet les quelques exemples fournis à ce sujet (Archæoptéryx, Ichtyostéga, Seymouria, Probainognathus, Homo habilis : toujours les mêmes !), qui sont habituellement allégués pour soutenir l’existence des « chaînons intermédiaires » sont en réalité tout à fait discutables ; alors que les hypothétiques ancêtres communs aux taxa supérieurs (ordres, classes, types) ne sont que des illusions, voire d’absurdes et grotesques chimères (figure 4). Archæoptéryx, l’oiseau jurassique centre-européen bien connu, ne peut représenter une passerelle solide entre reptiles et oiseaux, car ses prétendus « caractères reptiliens » (comme les dents dans le bec) ne sont en fait pas du tout « reptiliens » pour peu qu’on les examine de près (les tortues sont des reptiles, et pourtant elles n’ont pas de dents !).
On peut en dire autant de Probainognathus, un reptile cynodonte du Trias moyen (Argentine) qui, tout en présentant dans l’articulation cranio-mandibulaire certaines caractéristiques « de mammifère », ne peut pas être considéré comme un « chaînon intermédiaire » probant, puisque l’ensemble de ses caractères morphologiques le fait inexorablement tourner autour du « centre de gravité taxologique » des reptiles. Du reste, l’actuel ornithorynque d’Australie, s’il possède des caractéristiques « d’oiseau » (comme le bec semblable à celui du canard et la reproduction ovipare), n’en est pas moins, dans son ensemble, un véritable mammifère (figure 4).
Il faut d’ailleurs souligner avec force une dernière chose. Lorsqu’on récapitule l’histoire de la vie, du début du paléozoïque à aujourd’hui, l’augmentation progressive de la diversification systématique imaginée par Darwin (voir, par exemple, la seule figure reproduite dans L’origine des espèces) n’apparaît pas. La faune et la flore ont incontestablement changé d’une période à l’autre et, surtout, d’une ère à l’autre ; mais, somme toute, ces changements n’ont pas entraîné une complexification croissante. Il s’est agi, plutôt, de « variations sur des grands thèmes « restés inchangés depuis plus d’un demi-milliard d’années. Comme l’a également souligné le paléontologue David Raup [7], tout laisse à penser que le paléozoïque connut autant de taxa (espèces, genres, familles, ordres, classes et embranchements) que le mésozoïque, le cénozoïque et la période actuelle. Et cela suppose une condition fondamentale de « stabilité » qui est inconciliable avec l’idée évolutionniste dominante. Autrement dit, les flores et les faunes de l’ère paléozoïque n’étaient pas du tout moins complexes et moins diversifiées que les flores et faunes actuelles, mais simplement différentes, tout en restant fidèles aux mêmes plans fondamentaux d’organisation.
-

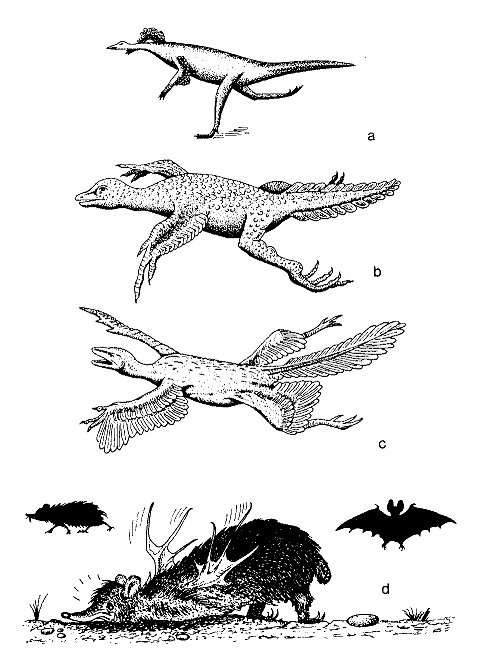
Figure 4
Illustration du caractère chimérique et mythique des « chaînons intermédiaires « ou « chaînons manquants ». Les trois premières figures qu’on observe de haut en bas se rapportent au fantomatique Proavis, le chaînon évolutif entre les Reptiles et les Oiseaux tant recherché, mais jamais découvert, par les paléontologues. Le baron von Nopcsa l’imaginait comme un petit reptile bipède et capable de courir qui, à force de sauts de plus en plus longs et de mouvements des « bras » de plus en plus rapides et fréquents, vit ses écailles se changer en plumes et se lança dans ses premiers vols. En revanche, Pycraft et Beebe se le représentaient comme une espèce de lézard arboricole qui commença à sauter d’une branche à l’autre, puis d’un arbre à l’autre, ses écailles se changeant dans le même temps en plumes. Il est clair que des créatures « intermédiaires » de ce genre, non encore capables de voler comme de vrais oiseaux et non plus capables de courir ou de ramper comme de vrais reptiles, n’auraient jamais pu vivre, car la sélection naturelle les aurait immédiatement éliminées. On peut en dire autant de bon nombre d’autres groupes animaux, à commencer par les Chiroptères, généralement considérés comme dérivés de petits Mammifères quadrupèdes comme les insectivores. Comment devons-nous donc imaginer les « chaînons manquants » ? Peut-être comme celui, vraiment chimérique et grotesque, de la figure humoristique (en bas) dessinée par Rutilio Sermonti ?
Peut-être aurez-vous tendance à estimer que la documentation paléontologique est trop maigre et inadéquate pour qu’on se permette de faire des généralisations, de quelque type que ce soit, sur le déroulement historique de la vie sur la Terre. Darwin lui-même était obligé de reconnaître que les objections les plus sérieuses contre sa théorie venaient précisément de la paléontologie ; mais il ne les jugeait pas décisives, ni même très significatives, puisqu’il estimait qu’elles étaient fondées sur des données très peu nombreuses et incertaines. Mais en réalité les choses ne se présentent pas ainsi. Bien que la fossilisation soit, en elle-même, un événement dépendant de conditions environnementales plutôt particulières, et donc assez rare, et bien que de nouvelles espèces de plantes et d’animaux fossiles soient sans cesse découvertes et décrites, chaque nouvelle acquisition n’invalide pas le cadre des connaissances précédentes, mais, simplement, le perfectionne et en précise les détails ; on peut même dire qu’elle en confirme et souligne constamment les connotations essentiellement discontinues. Les données statistiques de Alan B. Shaw [8] et les considérations de Chris Paul [9] sont à ce sujet plus qu’éloquentes, et nous autorisent à penser que la documentation paléontologique est parfaitement capable de nous fournir une description crédible — bien que toujours susceptible, évidemment, d’amélioration — de l’histoire de la vie sur notre planète. Je crois qu’il y a actuellement un consensus presque total parmi les paléontologues sur ce point fondamental. C’est pourquoi j’estime qu’on ne doit accorder aucun crédit à des affirmations dogmatiques et expéditives comme celles du célèbre ultradarwinien Ernst Mayr, qui a affirmé, dans un des derniers entretiens qu’il a accordés, que les paléontologues « ne sont pas du tout qualifiés pour discuter » [10] du problème de l’évolution. C’est exactement le contraire qui est vrai (figure 5).
|
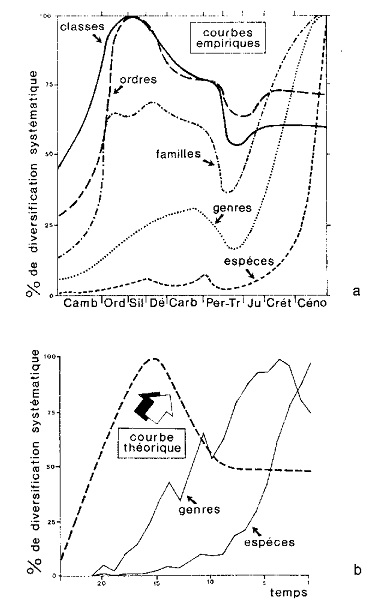
Figure 5 En haut : ce graphique montre la variation du nombre de taxa observée dans de nombreux groupes d’Invertébrés marins possédant des parties dures (donc susceptibles de s’être bien conservés à l’état fossile), du début de l’ère paléozoïque à la fin de l’ère cénozoïque. En ordonnées, les pourcentages de diversification pour chaque taxon (sont exclus les types, pour lesquels il n’y a eu aucune augmentation numérique des niveaux infra-paléozoïques aux niveaux cénozoïques). En abscisses, la succession des périodes géologiques. On voit que les taxa les plus élevés (classes et ordres) ont connu le maximum de diversité entre l’Ordovicien et le Silurien, avant d’entamer un brusque déclin jusqu’au Permien-Trias, à son tour suivi d’une stabilisation. Les familles ont atteint le maximum de diversité dans le Dévonien, subi un déclin à la fin de l’ère paléozoïque et enregistré une rapide remontée du Trias à la fin de l’ère cénozoïque. Les genres et surtout les espèces, en revanche, ont toujours connu un pourcentage croissant, de façon pratiquement constante, tout au long du temps géologique. Les variations de pourcentage des différents taxa doivent de toute façon être considérées comme apparentes et non réelles, dans la mesure où elles ne tiennent pas compte de plusieurs facteurs, dont notamment les différences d’étendue des affleurements rocheux (et donc aussi de la quantité d’espèces fossiles qu’on peut y trouver) relatifs à chaque période : étendue qui n’est pas du tout restée constante au fil du temps et qui a généralement augmenté des périodes les plus reculées aux périodes les plus récentes. En bas : le graphisme montre la simulation par ordinateur de la diversification dans le temps de 2000 espèces hypothétiques subdivisées en 100 genres, soit en ne tenant pas compte (lignes continues), soit en tenant compte (ligne pointillée, valable aussi bien pour les genres que pour les espèces) de la variation des surfaces d’affleurement rocheux relatives aux différentes périodes. La ligne pointillée correspond à celle des classes et des ordres du graphique précédent et semble donc s’appliquer indistinctement à tous les taxa. Bien que les deux graphiques (que nous avons empruntés à Raup) ne prennent en considération que les Invertébrés marins possédant des parties dures, ceux-ci, cependant, ne sont pas en nombre négligeable, ce qui mène à une conclusion nettement défavorable à l’évolutionnisme : la variété des formes de vie était élevée « dès l’origine » et est restée pratiquement constante à travers les périodes fossilifères. En d’autres termes, elle n ‘a pas augmenté progressivement dans le temps, comme le pensait Darwin. |
Il me semble que tous ces faits sont suffisants pour encourager la révision de certaines idées habituellement présentées comme allant de soi. Mais, quatre ans après la parution de Dopo Darwin, peut-on noter, au sein du monde scientifique, des signes de changement qui soient susceptibles de confirmer le contenu de votre livre ?
Je note avec satisfaction que depuis la sortie de notre livre les biologistes disposés à reconsidérer d’un œil critique les fondements de l’évolutionnisme orthodoxe sont de plus en plus nombreux. Il y a encore peu de temps, le seul fait de mettre en doute ces fondements était presque impensable et revenait à passer pour un partisan de l’obscurantisme et de la réaction. Aujourd’hui, cela se produit de moins en moins souvent. Cela prouve que Dopo Darwin n’est pas du tout le fruit d’une mentalité anachronique, aveuglée par des préjugés religieux — comme l’ont affirmé bruyamment certains —, mais le résultat de considérations et de réflexions sérieuses sur des problèmes importants que de nombreux autres scientifiques, indépendamment de nous et partout dans le monde, ne peuvent plus éviter de se poser. Il est clair que tout cela n’aurait pas lieu si l’évolutionnisme était vraiment capable de résoudre les principaux problèmes de la biologie.
Que la biologie s’éloigne de plus en plus du darwinisme orthodoxe, bien des faits le démontrent : les livres du zoologiste français Pierre-Paul Grassé [11] ; l’orientation « neutraliste « de l’école génético-moléculaire de Motoo Kimura et Masatoshi Nei [12] ; l’approche critique de généticiens de grand renom comme Richard Lewontin [13] et de paléontologues comme Chris Paul [14] ; le courant paléontologique des « équilibres ponctués « de Niles Eldredge, Stephen Jay Gould et Steven Stanley [15] ; le courant embryologico-épigénétique de Soren Lovtrup (qui a repris les thèses, jusque-là réputées hérétiques, de Goldschmidt, Schindewolf et Dalcq) [16] l’école systématique et cladistique de Willi Hennig [17], qui travaille dans le sillon tracé en son temps par notre compatriote Daniele Rosa [18] : l’école biogéographique de Gareth Nelson, Donn Eric Rosen et Norman Platnick [19], qui se fonde sur l’œuvre monumentale — jusqu’à maintenant délibérément ignorée — de Léon Croizat [20], lui-même ancien élève de Rosa ; le courant taxinomico-numérique de Philip Sneath et Robert Sokal [21], qui reprend — sans en développer cependant les aspects les plus positifs — les orientations théoriques et méthodologiques d’Alfredo Sacchetti [22]. On peut enfin citer, pour ce qui concerne notre pays, la Rivista di Biologia [23], dirigée par Giuseppe Sermonti, ouverte aux scientifiques anticonformistes, italiens et étrangers, et qui promet de devenir un bon creuset d’idées originales pour le renouvellement futur de la biologie.
Somme toute, il me semble donc que nous nous dirigeons vers une science libérée de la forme d’esprit darwinienne et de positions trop nettement conçues a priori, par conséquent vers une science ouverte à de nouvelles interprétations de l’énorme masse d’informations recueillies jusqu’à maintenant.
Vous souhaiteriez donc une espèce de pluralisme scientifique ?
Oui. Je souhaite surtout que chaque scientifique puisse dire librement ce qu’il pense et même avancer les thèses les plus audacieuses et les plus révolutionnaires, sans crainte d’être ostracisé dès le départ et mis à l’index, tout simplement parce quelques-uns de ses collègues particulièrement puissants et en mesure de faire la loi ne tolèrent pas ses idées.
_______________________________________________________
1 Giuseppe Sermonti et Roberto Fondi, Dopo Darwin. Critica all’evoluzionismo, Rusconi, Milano, 1982 (4ème édition).
2 Né en 1925, Giuseppe Sermonti a étudié la génétique des micro-organismes à l’Institut de la Santé de Rome, de 1950 1965. Il a publié Genetics of Antibiotic Producing Microorganismus (1969), Il crepuscolo dello scientismo (1971), Genetica Generale (1971 et 1974), La mela di Adamo e la mela di Newton (1974), Le forme della vita (1981). Depuis 1974, Giuseppe Sermonti est professeur de génétique à l’université de Pérouse, où il dirige l’Institut d’Histologie et d’Embryologie. Il s’est rendu à Moscou en 1978 en tant que vice-président du XIVème Congrès international de génétique. Le professeur Sermonti dirige, depuis la fin de l’année 1979, la Rivista di Biologia (N.D.T.).
3 Pierre-Paul Grassé, L’évolution du vivant, Albin Michel, 1973.
4 Fred Hoyle et Chandra Wickramasinghe, Le nuage de la vie. Les origines de la vie dans l’univers, Albin Michel, 1980.
5 Francis Crick, La vie vient de l’espace, Hachette, 1982.
6 Alfredo Sacchetti et Roberto Fondi, Verso una tassologia del Systema Naturae –Towards a Taxology of the « Systerna Naturae », in Rivista di Biologia, 75, 2, 1982, pp. 273-292.
7 David M. Raup, Taxonomic Diversity during the Phanerozoic, in Science, 177, 1972.
8 Alan B. Shaw, Time in Stratigraphy, McGraw-Hill, New York, 1964.
9 Chris Paul, The Natural History of Fossils, Weidenfeld and Nicolson, London, 1980.
10 In Omni, mai 1983.
11 Cf. Pierre-Paul Grassé, Op. cit., 1973 ; Toi, ce petit Dieu, Albin Michel, 1971.
12 Motoo Kimura, La teoria della neutralità nell’evoluzione molecolare, in Le Scienze, 137, janvier 1980 ; Masatoshi Nei, Molecular Population Genetics and Evolution, North-Holland Publications Company, Amsterdam-Oxford, 1975.
13 Richard C. Lewontin, The Genetic Basis of the Evolutionary Change, Columbia University Press, New York, 1974.
14 Chris Paul, Op. cit., 1980.
15 Cf. Niles Eldredge et Stephen Jay Gould, Punctuated Equilibria : an Alternative to Phyletic Gradualism, in Schopf, Freeman, Cooper et al. (ed.), Models of Paleobiology, San Francisco, 1972 ; Steven M. Stanley, The New Evolutionary Timetable, Basic Books, New York, 1981.
16 Cf. Soren Lovtrup, Epigenetics : a Treatise on Theoretical Biology, Wiley, London, 1974.
17 Willi Hennig, Grundzüge einer Theorie der phylogenetischen Systematik, Deutscher Zen-tralverlag, Berlin, 1950 (tr. anglaise : Phylogenetic Systematics, University of Illinois Press, Urbana, 1966).
18 Daniele Rosa, L’Ologenèse, Félix Alcan, 1931.
19 Cf. Gareth Nelson et Donn E. Rosen, Vicariance Biogeography. A Critique, Columbia University Press, New York, 1981 ; Gareth Nelson et Norman Platnick, Systematics and Biogeography : Cladistics and Vicariance, Columbia University Press, New York, 1981.
20 Léon Croizat, Space, Time, Form : the Biological Synthesis, publié par l’auteur, Caracas, 1962. Voir également : Léon Croizat, Gareth Nelson et Donn E. Rosen, Centers of Origin and Related Concepts, in Systematic Zoology, 23, 1974.
21 Philip H.A. Sneath et Robert R. Sokal, Numerical Taxonomy, Freeman, San Francisco, 1973.
22 S. Beer et Alfredo Sacchetti, Problemi di sistematica biologica, Einaudi, Torino, 1952.
23 La revue est publiée par l’Institut de Biologie de l’université de Pérouse (Rivista di Biologia — Casella Postale 317 — 06100 Perugia).