
Dans tout ce que vous avez affirmé, je crois voir transparaître l’un des aspects classiques propres à une vision alternative de la réalité scientifique, à savoir la délimitation des problèmes à approfondir et l’abandon des champs de recherche précédemment étudiés et désormais réputés privés de sens, «non scientifiques». Comme l’a souligné Thomas Kuhn [1], l’histoire de la science nous présente ces phénomènes lorsque de nouveaux «paradigmes» remplacent, ou tendent à remplacer, les anciens paradigmes. Mais si pour expliquer scientifiquement le phénomène vivant une logique de type évolutionniste et déterministe ne suffit pas, s’il faut aussi une logique de type cybernétique, quelle devrait être, alors, la philosophie de base y correspondant ? Quel paradigme alternatif les biologistes devraient-ils adopter, selon vous, à partir de maintenant ?
Personnellement, je n’ai guère de doutes à ce sujet. Puisque la seule clef de lecture des systèmes cybernétiques consiste, comme je l’ai déjà dit, à les considérer non comme des mécanismes réglés par des processus purement déterministes, mais comme des totalités organiques douées de «programme informant» (dont le développement spatio-temporel est un processus organisé, typiquement anti-stochastique, qui corrige erreurs et déviances par référence constante à ce «programme», et qui ne converge pas vers un état-limite plus probable que la situation initiale, mais atteint au contraire, à travers des mémorisations et élaborations informelles, un état plus ordonné capable d’autorégulation, d’adaptation, d’homéostase et d’autodirection), la philosophie de base de la biologie ne pourra être que la philosophie organiciste. Cette dernière, en effet, repose précisément sur le principe dit «holiste», déjà clairement formulé par Aristote, et selon lequel l’organisme est un tout, un tout qui est plus que la simple somme de ses parties. Ce qui caractérise l’organicisme par rapport aux conceptions mécanistes et sommatives, selon lesquelles le tout correspond — ni plus ni moins — à la simple résultante des parties, c’est justement l’affirmation de l’unité, de la totalité, de l’individualité, de l’organisation et de la nouveauté de l’organisme vivant.
Que les organismes vivants se comportent aussi comme des machines — en tant qu’ils comprennent des structures (comme les os, les muscles, les vaisseaux sanguins, etc.) qui fonctionnent comme de véritables machines (ex.: leviers, tirants, tubes, filtres, etc.) — est un fait. Ce fait, cependant, ne signifie pas que les vivants sont des machines.
Il y a des différences fondamentales entre les machines et les organismes. Les machines sont construites, les organismes croissent. Les activités d’une machine résultent exclusivement de sa structure, alors que chez les organismes la structure résulte des activités de ses processus. Les machines sont construites parce qu’on met ensemble un nombre bien défini de parties, de manière précise et préétablie, tandis que les organismes — tout en présentant des morphologies et des types de comportement bien définis — peuvent s’offrir un degré parfois très élevé de flexibilité et de plasticité (au point qu’il n’y a pas deux organismes qui aient des parties totalement identiques). Les machines fonctionnent selon des chaînes linéaires de cause à effet, si bien que lorsqu’elles tombent en panne, on peut habituellement découvrir une seule cause responsable de la panne. Au contraire, le fonctionnement des organismes est dirigé par des flux circulaires d’information connus comme cycles de feed-back ou de rétroaction. Par exemple, la partie A peut influencer la partie B ; B peut influencer C ; et C peut rétroagir sur A, fermant ainsi harmonieusement le cycle. En général, dans un système de ce type la perturbation est due à de multiples facteurs, qui peuvent s’amplifier réciproquement à travers des cycles interdépendants de rétroaction. Cette interconnexion non linéaire des organismes vivants ne rend pas seulement problématiques les tentatives de la médecine classique pour associer les maladies à des causes particulières ; elle montre aussi — entre autres choses — le caractère illusoire du «dogme central» de la biologie, c’est-à-dire la croyance selon laquelle les différentes structures (physiques ou mentales) d’un organisme individuel sont contrôlées ou «dictées» de manière univoque par son ADN. La vision organiciste, en effet, permet de comprendre clairement que l’ADN ne détermine pas irréversiblement le fonctionnement d’un organisme comme les ressorts et les roues dentelées déterminent celui d’une montre, mais fait partie intégrante d’un tout et doit donc se conformer à l’organisation systémique de ce tout.
Cela veut-il dire que ce que soutient la sociobiologie [2] la plus extrémiste, à savoir que le comportement des vivants se justifierait en fonction d’une meilleure préservation et d’une meilleure «administration» de leur ADN au cours du conflit impitoyable pour l’existence, est faux ?
Je ne dirais pas que cela est faux, mais que cela n’est vrai que pour moitié. Il faudrait dire en effet que l’ADN existe aussi pour être au service des vivants et de leur comportement. Quel biologiste pourrait être en mesure de démontrer que cette seconde alternative n’est pas légitime ?
Poursuivons.
En conclusion, un organisme vivant est donc un système qui s’auto-organise, dont l’ordre structurel et fonctionnel ne peut pas être simplement imposé par l’environnement, mais qui est établi par lui-même. Ainsi l’organisme ne correspond-il pas seulement à la machine, mais aussi au projet ou pensée inventive qui la justifie. Cela était très clair pour Aristote également, qui percevait un principe téléologique au sein de l’être naturel, sans le rapporter à un Dieu ordonnateur «extérieur», mais qui le jugeait inhérent à l’être naturel lui-même, et indissolublement associé au principe de causalité ordinaire.
Je suis convaincu que c’est là la façon la plus correcte d’interpréter le phénomène vivant, et même l’univers réel dans sa totalité. Ludwig von Bertalanffy ne pensait pas autrement, lui qui écrivait dans sa Théorie générale des systèmes : «Nous ‘voyons’ actuellement l’univers comme une énorme hiérarchie, depuis les particules élémentaires aux noyaux atomiques, aux atomes, aux molécules, aux composés moléculaires, à l’abondance des structures (électrons et particules lumineuses) entre les molécules et les cellules, aux cellules, aux organismes et au-delà, aux organisations supra-individuelles (les taxa, N.D.L.A.) (…) On trouve une hiérarchie semblable à la fois dans les ‘structures’ et dans les ‘fonctions’. En dernier ressort la structure (c’est-à-dire l’ordre des parties) et la fonction (l’ordre des processus) sont peut-être la même chose : dans le monde physique la matière se dissout en un jeu d’énergies et dans le monde biologique les structures expriment un flux de processus» [3].
Plus récemment, Fritjof Capra, dont nous avons déjà parlé, a écrit des pages très significatives à ce sujet, et qui s’accordent parfaitement avec l’esprit de von Bertalanffy : «Une compréhension plus complète de la vie — écrit Capra — ne sera atteinte que si l’on développe une ‘biologie des systèmes’, une biologie qui voit l’organisme comme un système vivant, plutôt que comme une machine. La vision systémique considère le monde en termes de relations et d’intégration. Les systèmes sont des touts intégrés dont les propriétés ne peuvent être réduites à celles de plus petites unités. Au lieu de se concentrer sur les éléments fondamentaux ou sur les substances de base, l’approche systémique met l’accent sur les principes de l’organisation. Les exemples de systèmes abondent dans la nature. Chaque organisme — de la plus infime bactérie jusqu’aux humains en passant par toute la variété végétale et animale — est un tout intégré et, donc, un système vivant. Les cellules sont des systèmes vivants, de même que les tissus et les organes du corps, le cerveau humain en étant l’exemple le plus complexe. Mais les systèmes ne sont pas l’apanage des organismes individuels et de leurs parties. Les mêmes aspects d’intégralité sont observables dans les systèmes sociaux — tels qu’une fourmilière, une ruche ou une famille humaine — et les écosystèmes qui sont formés d’un ensemble d’organismes et de matière inanimée en interaction mutuelle (…) La majorité des organismes ne sont pas simplement inclus dans les écosystèmes, ils sont des écosystèmes eux-mêmes contenant un ensemble d’organismes plus petits jouissant d’une autonomie considérable mais ne s’en intégrant pas moins harmonieusement au fonctionnement du tout (…) La tendance qu’ont les systèmes vivants à former des structures à plusieurs niveaux, chaque niveau différant par sa complexité, est omniprésente dans la nature et doit être considérée comme un principe fondamental de l’auto-organisation (…) Une cellule peut faire partie d’un tissu, mais peut également être un microorganisme faisant partie d’un écosystème et, bien souvent, il est impossible de dessiner une frontière précise entre ces descriptions. Chaque sous-système est, à la fois, un organisme relativement autonome et un composant d’un organisme plus vaste ; c’est un ‘holon’, pour reprendre le terme d’Arthur Koestler, manifestant les propriétés indépendantes d’un tout et les propriétés dépendantes d’une partie» [4].
Capra en arrive donc aux mêmes conclusions que von Bertalanffy, Lovelock et Margulis : la Terre et même toute la nature ne peuvent être correctement comprises que si on les considère comme des organismes vivants.
Oui. Et c’est pourquoi on songe alors spontanément aux auteurs qui conçoivent l’univers non pas seulement comme un immense organisme vivant auto-organisé, mais aussi comme une entité consciente. Nous pouvons nous référer, par exemple, aux scientifiques de la «gnose de Princeton», pour reprendre l’expression de Raymond Ruyer [5] (Gustav Stromberg, Victor F. Weisskopf, E.T. Whittaker, Carl Friedrich von Weizsäcker, G.J. Withrow, Dennis W. Sciama, David Bohm, R.P. Feynman, Walter M. Elsasser, E.P. Wigner, etc.), au cybernéticien David Foster [6] et surtout à l’anthropologue Gregory Bateson.
L’auteur de La nature et la pensée [7] ?
En effet. Bateson estime que la pensée est une propriété essentielle des systèmes vivants. Déjà dans les années quarante, le biologiste George Coghill était parvenu à des conclusions semblables. Il distinguait en effet dans les organismes vivants trois types d’organisation en interaction étroite : structure, fonction et «mentation» (mentation). La structure correspondrait à l’organisation dans l’espace, la fonction à l’organisation dans le temps et la mentation à un type d’organisation qui intégrerait les deux précédentes à un niveau plus élevé de complexité. Elle correspondrait, en somme, à l’organisation de toutes les structures-fonctions et serait donc, en même temps, une métastructure et une métafonction. Dans les systèmes vivants de complexité faible, elle représenterait ce qu’on appelle habituellement «comportement» (behaviour), tandis que dans les systèmes plus élevés et chez l’homme elle présenterait aussi les caractéristiques de non-spatialité et de non-temporalité qui sont généralement associées à l’idée d’«esprit».
Puisque, selon Bateson, cette mentation correspond aux lois d’organisation des systèmes vivants, et puisque ces derniers ne comprennent pas seulement des organismes individuels, mais aussi des formes sociales (les taxa) et écologiques, il s’ensuit qu’elle est immanente à l’homme, mais aussi à tout le système écologico-social dont nous constituons autant de sous-systèmes. Ce qui a de très vastes implications culturelles, car cela mène à la conclusion que, de même qu’il existe toute une hiérarchie de structures vivantes, de même il existe une hiérarchie correspondante de structures mentales ou psychiques. Il y aura, par exemple, des niveaux de mentation «métabolique» organisant les structures et les fonctions des cellules, des tissus et des organes ; et des niveaux de mentation «spécifique», «générique», «familiale», organisant les morphologies spatio-temporelles des diverses espèces, genres, familles, etc. Et tous ces niveaux, sujets à leur logica minor particulière, feront à leur tour partie intégrante de la logica major inhérente à la totalité du cosmos.
«Dans l’ordre stratifié de la nature, écrit encore Capra, les esprits humains individuels sont intégrés dans les esprits plus vastes des systèmes sociaux et écologiques, et ceux-ci sont, à leur tour, intégrés dans le système mental planétaire — l’esprit de Gaïa — qui, lui aussi, doit participer à une sorte d’esprit universel ou cosmique. Le cadre conceptuel de la nouvelle approche systémique n’est en aucune manière restreint par une éventuelle association entre cet esprit cosmique et l’idée traditionnelle de Dieu. Selon Jantsch : ‘Dieu n’est pas le créateur mais l’esprit de l’univers’. Vue sous cet angle, la déité n’est ni mâle ni femelle, elle n’est pas non plus manifeste sous une forme personnelle ; elle ne représente ni plus ni moins que la dynamique auto-organisatrice de l’ensemble du cosmos» [8].
La conception de l’Être comme kosmos ou Systema Naturae, c’est-à-dire comme ordre hiérarchique impliquant des niveaux multiples à complexité variable, en interaction et interdépendance mutuelles telles qu’ils expriment une intelligence supérieure, a toujours joué un rôle prépondérant dans les traditions non modernes de tous les peuples. Cette conception s’est maintenue régulièrement jusqu’à Linné et Cuvier, et seule la diffusion du mythe évolutionniste a pu l’éloigner des esprits des scientifiques. Aujourd’hui, cependant, cette conception fait retour, inexorablement. Et peu importe que cela se produise à travers une grande contradiction. Le fait demeure, et c’est cela qui importe le plus.
II y a peu, il était fait allusion, dans une phrase de Capra citée par vous, à la notion de holon, développée et proposée par Koestler, notamment dans son livre intitulé Janus [9]. Puisque ce livre est aussi un résumé des idées soutenues par son auteur dans le domaine des sciences de la vie, je suis curieux de savoir ce que vous en pensez.
Koestler était un excellent écrivain, un homme courageux et très cultivé, et c’est pourquoi on le lit toujours avec plaisir. Toutefois, il faut le lire avec esprit critique, dans la mesure où, bien qu’il adopte souvent des points de vue très proches de ceux soutenus ici, il tombe lui aussi, parfois, dans des contradictions flagrantes.
Dans le livre que vous citez, Koestler considère chaque réalité vivante comme un «holon», c’est-à-dire comme un tout organique assimilable à une sorte de Janus bifrons horizontalement déployé, qui regarde simultanément vers le haut (les niveaux ou holons supérieurs dont il est partie intégrante) et vers le bas (les niveaux ou holons inférieurs dont il est lui-même composé). Bien que le terme de holon n’ait qu’une valeur relative (les holons composant un holon de niveau plus élevé devraient, en réalité, être appelés mérons par rapport à ce dernier), on peut certainement l’accepter. Ce qui n’est pas acceptable, en revanche, ce sont certains points de vue pour le moins aberrants de Koestler, comme par exemple celui relatif au conflit qui existerait, dans l’espèce humaine, entre le cortex cérébral ou néocortex (typiquement humain en tant que siège responsable de l’intelligence et du raisonnement) et la structure encéphalique plus profonde ou paléocortex, d’origine «reptilienne» (responsable des activités instinctives et endocrino-végétatives). Selon Koestler — qui n’a certes pas une image flatteuse de l’homme —, les comportements agressifs et violemment antisociaux manifestés par notre espèce ne seraient que le résultat d’une «erreur évolutive» consistant précisément dans le fait que la superposition du néocortex au paléocortex serait arrivée trop rapidement et trop brutalement pour permettre une coordination harmonieuse et contrôlée entre l’un et l’autre. Cette erreur évolutive — associée à une autre, due au fait que les petits de l’homme doivent supporter une période d’impuissance et de dépendance totales par rapport à leurs parents plus prolongée que celle des petits de n’importe quelle autre espèce — pourrait être corrigée et neutralisée, selon Koestler, au moyen de doses répétées de médicaments à base de combinaisons enzymatiques ; et il faudrait faire cela rapidement, pour éviter que l’humanité ne finisse par s’autodétruire à brève échéance.
Ces idées (qui reprennent en partie des thèses déjà soutenues par des auteurs comme Paul McLean, Luigi Valzelli et le vulgarisateur scientifique Carl Sagan) me paraissent, pour être tout à fait franc, assez ridicules, car le néocortex n’est certainement pas quelque chose qui se développe indépendamment du reste du système nerveux. Tout le développement des opérations cérébrales se ressent constamment des signaux qui parviennent au néocortex depuis les secteurs les plus profonds et dépend de la façon dont le néocortex lui-même élabore, contrôle et utilise les contenus de ces signaux. Si nous tenons compte de cela, il est clair, alors, que nous ne pouvons pas attribuer la responsabilité de toute conduite antisociale, violente et agressive (Koestler l’appelle même «schizophysiologique») au «cerveau reptilien», puisque le cortex cérébral contribue, de façon inévitable et responsable, à cette conduite. Ces deux cerveaux, en somme, ne sont pas des mondes séparés, ils sont complémentaires, en interrelation permanente, si bien que le comportement humain ne saurait être que la résultante de leur interaction. Il est vraiment surprenant qu’un auteur comme Koestler — partisan ardent d’une vision non réductionniste, mais «systémique», de la réalité, comme von Bertalanffy et Capra — finisse par tomber dans des erreurs de ce genre.
Quant au fait que l’homme vient au monde dans une phase de développement beaucoup plus précoce que celle de tous les autres animaux — phase qui selon Koestler frustrerait le nouveau-né et contribuerait dès le départ à le maintenir dans un état névrotique permanent, dans la mesure où elle le rendrait pour très longtemps totalement dépendant de ses parents ou d’autres êtres humains adultes («Le lavage de cerveau commence au berceau», dit-il [10]) — on ne voit vraiment pas quelle autre alternative Koestler est capable de proposer. Comme l’ont bien montré Adolf Portmann [11] et Arnold Gehlen [12], la phase de développement précoce a une importance fondamentale chez l’homme, car c’est elle qui lui fait acquérir la station debout, apprendre le langage proprement dit et recevoir l’habitude à la pensée et à l’action techniques. Abandonnons un nouveau-né à lui-même, et il mourra très vite. Éloignons-le des hommes pour le confier à des loups, des singes, etc., et il ne deviendra certainement pas un Romulus, un Mowgli ou un Tarzan, mais un enfant destiné à vivre très peu de temps et dans des conditions semi-animalesques bien plus tarées — sur le plan psychologique — que la condition que Koestler assigne à l’humanité «normale».
Revenons à l’idée d’ordre hiérarchique ou stratifié de la réalité. Je voudrais saisir plus clairement comment vous l’entendez. Bien qu’il corresponde au Systema Naturae de Cuvier (qui présentait les mêmes caractéristiques fondamentales que celui de Linné, à ceci près qu’il reconnaissait les changements «révolutionnaires» subis par les flores et les faunes au cours de l’histoire géologique), il me semble que cet ordre ne peut pas être considéré comme une structure rigidement statique, mais doit admettre pour le moins un certain dynamisme dans l’espace et dans le temps. Comment concevez-vous ce dynamisme ? Ne pourrait-il pas coïncider avec celui des «transformistes finalistes» ? En somme, ne pourrait-il pas rentrer lui aussi, ne serait-ce que dans ses grandes lignes, dans le cadre du paradigme évolutionniste ?
Il est clair que je ne conçois pas du tout le Systema Naturae comme une structure rigidement statique. Du reste, ni Linné ni Cuvier ne le concevaient ainsi. Ils admettaient la variation et le mouvement ; mais ils ne les considéraient pas — contrairement à Buffon et aux modernes «visionnaires évolutifs» — comme des manifestations d’un pouvoir créateur débridé et inépuisable de la nature, mais comme des phénomènes contrôlés et maintenus à l’intérieur de limites spécifiques. Dans ce tourbillon de mouvement et de variabilité, les formes des vivants — numériquement très élevées mais pouvant être ramenées, dans leur ensemble, à quelques plans structurels fondamentaux seulement (quatre selon Cuvier et environ deux douzaines selon les biologistes actuels, dans le seul règne animal) — restaient relativement constantes, ne changeant qu’au cours de leur cycle de naissance, de croissance, de reproduction et de mort. Pour Linné et Cuvier, ce mouvement vital auto-perpétué était tout à fait différent des mouvements ordinaires de la matière. La vie était pour eux un système de processus menant à l’expression et au maintien de certains modèles structurels ; et ceux-ci, à leur tour, étaient construits et coordonnés entre eux de manière à activer les processus essentiels aux activités inhérentes au Système lui-même. Ce Systema Naturae est donc une structure dynamiquement statique, avec un dynamisme qui ne se déroule pas de façon linéaire et indéfinie, mais bien dans le cadre de «cercles de formes» (pour reprendre une expression du zoologiste allemand Otto Kleinschmidt, qui vécut à la fin du XIXème siècle et au début du XXème) spécifiques comparables aux holons de Koestler. Cela revient aussi à reconnaître que la hiérarchie de holons dont se compose la réalité biologique n’est pas constituée seulement de formes au sens spatial, mais aussi de «formes temporelles», lesquelles, étant cycliques, sont justement tout aussi bien définies que les premières : de sorte que chaque cycle temporel particulier d’un holon donné s’intégrera dans le cycle temporel du holon de niveau supérieur dont le premier est une partie inséparable.
Capra, encore lui, a écrit à ce sujet des choses vraiment illuminantes : «Les modèles rythmiques semblent présents à tous les niveaux. Les atomes sont des modèles d’ondes de probabilité, les molécules sont des structures vibrantes et les organismes sont des modèles de fluctuations multidimensionnels, interdépendants. Les plantes, les animaux et les êtres humains traversent des cycles d’activité et de repos et toutes leurs fonctions physiologiques oscillent suivant des rythmes à périodicités diverses. Les composants des écosystèmes sont reliés par des échanges cycliques de matière et d’énergie ; les civilisations se développent puis déclinent en cycles évolutifs, et la planète dans son ensemble a ses rythmes et ses récurrences alors qu’elle tourne autour de son axe et du soleil. Les modèles rythmiques sont donc un phénomène universel, mais n’en permettent pas moins aux individus d’exprimer leurs personnalités distinctives. L’expression d’une identité personnelle unique est une caractéristique importante des êtres humains, et il semble que cette identité pourrait être, essentiellement, une identité de rythme. On peut reconnaître les individus humains à leur façon de parler caractéristique, aux mouvements de leur corps, à leur gestuelle, à leur façon de respirer ; autant d’éléments représentant différents types de modèles rythmiques. De plus, il est de nombreux rythmes ‘figés’, comme nos empreintes ou notre écriture, qui sont également uniques aux individus (…) Deux thèmes fondamentaux reviennent constamment dans l’étude de la matière vivante et inorganique (…) l’interconnexion et l’interdépendance universelles de tous les phénomènes, et la nature intrinsèquement dynamique de la réalité» [13].
La considération du métabolisme organique, qui comporte des rythmes contrôlés et coordonnés de production et de destruction cellulaires, nous indique aussi quelle est la perspective la plus appropriée pour interpréter le phénomène de la mort : «Nous avons vu que l’auto-régénération — la destruction suivie de la reconstruction de structures en cycles continus — est un aspect essentiel des systèmes vivants. Mais les structures qui sont continuellement remplacées sont elles-mêmes des systèmes vivants. Dans cet ordre d’idées, l’auto-régénération du système plus large est son propre cycle de naissance et de mort. La naissance et la mort apparaissent donc, maintenant, comme constituant un aspect central de l’auto-organisation, l’essence même de la vie (…) Puisque nous aussi sommes nés et destinés à mourir, cela signifie-t-il que nous faisons partie de systèmes plus vastes qui se renouvellent constamment ? Cela semble bien être le cas. Comme toutes les autres créatures vivantes, nous appartenons à des écosystèmes et nous formons également nos propres systèmes sociaux. Finalement, à un niveau beaucoup plus large, il y a la biosphère, l’écosystème de toute la planète, dont notre survie dépend totalement» [14].
Je crois que cela suffit à faire comprendre en quel sens je me déclare organiciste. De même que chaque forme vivante représente une totalité spatio-temporelle auto-organisée, en tant qu’elle reste fidèle à sa morphologie spatiale particulière et à sa dynamique spécifique de nature cyclique ou rythmique (naissance, croissance, déclin et mort), de même toute la nature constitue une totalité spatio-temporelle structurée en niveaux multiples à complexité variable et incluant le temps (avec tous les événements passés, présents et futurs). Ma conception unitaire n’a en commun avec le «transformisme finaliste» que la reconnaissance du caractère dynamique du Systema Naturae, dynamisme qui est vu comme se déployant selon les fins inhérentes aux lois architecturales du système lui-même, et donc comme le déroulement d’un plan [15] . Mais pour tout le reste, elle en diffère radicalement, car :
1) elle ne postule pas au départ ce qui devrait être démontré, mais justifie la finalité naturelle sur la base d’argumentations scientifiques sûres et contrôlables, fondées essentiellement sur la notion de syntropie et sur la théorie des modèles d’univers de Fantappié ;
2) elle ne s’intéresse pas à ce qu’on appelle les «arbres phylogénétiques», auxquels je ne crois pas et dont la recherche m’apparaît comme une perte de temps pure et simple ;
3) elle interprète le monde des vivants comme un système hiérarchique unitaire de «cercles de formes» spatio-temporels.
Tout cela coïncide parfaitement, dans ses grandes lignes, avec les visions de la nature d’Aristote, Linné et Cuvier : visions qui sont classiquement présentées comme des exemples de pensée franchement «fixiste».
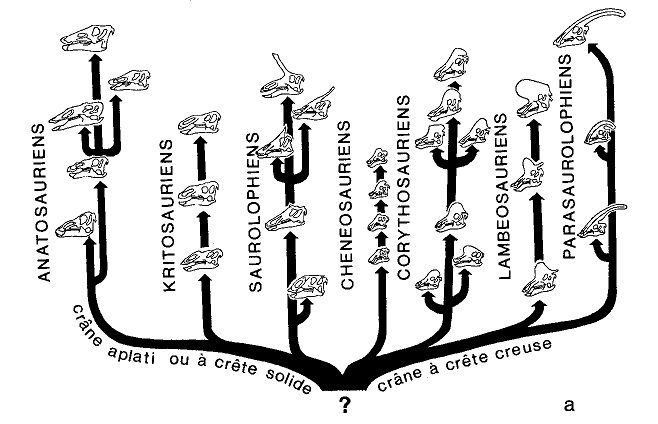
Je vais maintenant vous poser une question très concrète. J’ai ici avec moi une figure extraite d’un livre sur les Dinosaures écrit par l’évolutionniste Beverly Halstead (figure 7) [16]. La figure représente l’arbre phylogénétique des Hadrosauriens ou Dinosaures «à bec de canard». Si je décidais d’adhérer à votre point de vue, comment devrais-je modifier cette figure ?
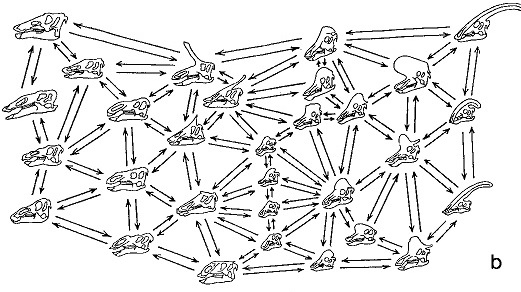
En la complétant, tout simplement. Au lieu d’avoir seulement des lignes avec des flèches à sens unique selon la direction du temps géologique, il faudrait placer des flèches à double sens dans toutes les directions (figure 7). Cela pour signifier que chaque taxon ne dépend pas (ou n’est pas causé) seulement par un seul taxon antérieur à lui, mais aussi par les autres taxa antérieurs, par les taxa contemporains et par les taxa à venir. De mon point de vue, en effet, l’ensemble des Hadrosauriens n’est pas un simple agrégat mécanique d’éléments reliés entre eux par des rapports de caractère déterministe et linéaire, mais un système hautement organisé de «holons» en interrelation étroite et réciproque.
|
Figure 7a Interprétation scientifique conventionnelle (évolutionniste-darwinienne) d’un complexe organique quelconque, en l’occurrence celui des Dinosaures à bec de canard ou Hadrosauriens (d’après Halstead). L’explication adhère de façon stricte aux postulats de la physique classique : elle voit chaque forme vivante comme causée de manière univoque et déterministe par une forme antérieure. Le tout, par ailleurs, sera déterminé par une seule cause initiale, consistant dans la fameuse «espèce ancestrale» ou «chaînon manquant», qui reste toujours indéfinissable et inexistante dans la documentation fossile concrète.
Interprétation scientifique du même groupe animal en fonction du paradigme organiciste. Ici, chaque forme vivante est vue comme partie intégrante d’un système spatio-temporel de nature cybernétique (ou informationnelle), et non plus simplement de nature déterministe, au sens classique et galiléen du terme. Des processus de feed-back ou d’interaction réciproque lient étroitement entre elles les formes ou éléments du système, et ce indépendamment du fait qu’elles soient contemporaines ou qu’elles existent en des temps différents ; elles occupent simplement des positions différentes dans le même bloc spatio-temporel ou chronotope biologico-naturel. Sur la base de la théorie moderne de la relativité, en effet, aussi puisque le présent existe avec ce que nous appelons «passé» et «futur», les choses passées, futures, conditionnent et «causent» le présent. |
Mais de quelle façon peut-on évaluer sous une forme objective, vraiment scientifique, la nature de ces interrelations ?
Je pense qu’aujourd’hui, grâce aux progrès des sciences mathématiques et techniques (je songe notamment aux ordinateurs), cela peut être fait de différentes façons, même si, pour le moment, je n’en ai qu’une en tête, de type nettement systématique.
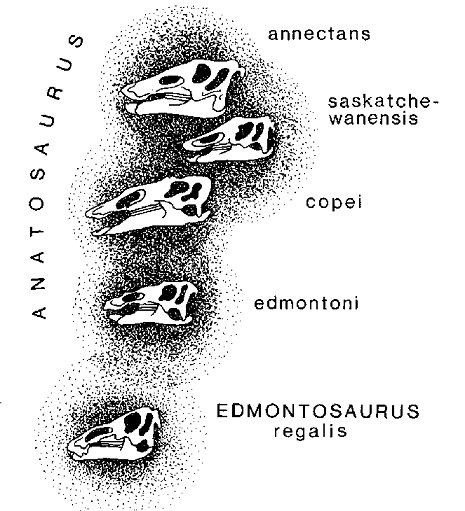
Pour simplifier, prenons seulement une partie du système des Hadrosauriens, à savoir le sous-système des Anatosauriens. Je crois qu’il est permis et utile de considérer les taxa qui intègrent ce sous-système comme les «entités» étudiées par la physique contemporaine, donc comme des nuages de probabilités d’interrelation (ou, pour reprendre une expression de l’anthropologue Alfredo Sacchetti, d’«adhérence typologique»). Si nous représentons ces nuages par des ensembles de points (les individus composant les taxa), on verra alors qu’à des probabilités élevées d’interrelation correspondront de fortes superpositions entre les nuages (figure 8). Le cas-limite de probabilité maximale, égale à l’unité, on l’aura lorsque les nuages finiront par «adhérer» complètement entre eux. Dans ce cas, les taxa qu’ils représentent seront considérés comme identiques. Inversement, plus les superpositions entre les nuages tendront à être petites, plus les taxa seront considérés comme différents et systématiquement éloignés entre eux.
|
Fig 8 Constitués de nombreux éléments semblables entre eux, les différents systèmes biologiques (en l’occurrence celui des Hadrosauriens) peuvent être considérés — conformément à ce qu’on fait pour étudier le monde subatomique — comme des «nuages de probabilités d’interaction» (ou, pour reprendre l’expression de l’anthropologue Sacchetti, d’«adhérence typologique») avec d’autres systèmes plus ou moins proches. Puisque ces probabilités peuvent être déterminées grâce à l’outil statistique et mathématique, la possibilité se présente de parvenir à une véritable science biosystématique, donc à une taxologie (et non plus à une simple «taxonomie» ayant un fondement purement conventionnel : souvenons-nous de la distinction entre les termes grecs logos et nomos, désignant respectivement la loi de la réalité naturelle et celle créée par l’homme pour ses seuls besoins pratiques). Les systèmes biologiques composés d’éléments à interaction forte seront constitutionnellement homogènes et situés par conséquent très bas dans l’échelle hiérarchique du Systema Naturae (ex.: races, espèces, genres), alors que les systèmes composés d’éléments à interaction faible seront constitutionnellement différenciés et situés par conséquent sur des positions élevées de la même échelle (ex.: ordres, classes, types). À une «adhérence typologique» de valeur limitée correspondra une discontinuité statistique au sein de la totalité spatio-temporelle dont cette adhérence, ou interaction, est partie intégrante. |
Il est fondamental de comprendre que les superpositions pourront tendre vers zéro tant qu’on voudra, mais ne parviendront jamais à s’annuler complètement. À l’intérieur du Systema Naturae, en effet, tout est relié (autrement, parler de système n’aurait pas de sens), et chaque distinction que nous pourrons y faire n’aura évidemment de valeur que dans un sens probabiliste.
Je crains de n’avoir pas bien compris votre dernière affirmation.
Je chercherai à m’expliquer plus clairement. Puisque les taxa biologiques sont des entités collectives, formées de populations, il est évident que pour les étudier scientifiquement, nous devons surtout recourir à des instruments logiques de type statistique, c’est-à-dire probabiliste. Essayons alors d’établir la relation existant entre la longueur du membre postérieur de l’homme et celui de la grenouille. Une fois que sera disponible un nombre convenable de mesures de cette longueur pour les deux espèces, il suffira de dessiner sur le même graphique les courbes de fréquence correspondantes.
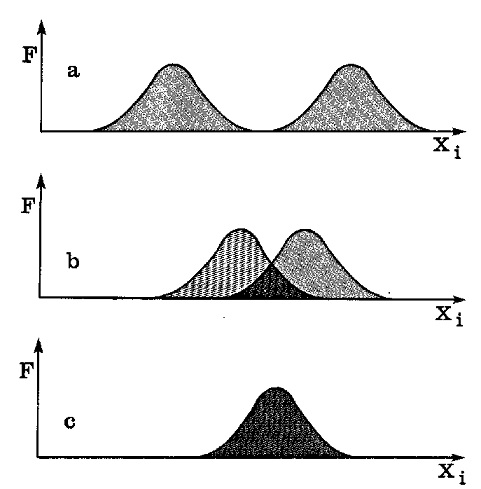
On sait que les courbes de fréquence, expression matérielle de courbes de probabilité, sont, comme celles-ci, asymptotiques. La probabilité de transvariation (ou de superposition) entre les courbes pourra donc devenir pratiquement nulle comme dans le cas de l’homme et de la grenouille, mais ne se réduira jamais absolument à zéro (figure 9).
On peut dire que pour évaluer la quantité de la «probabilité d’interrelation» entre les groupes de vivants les plus variés, il faudrait en somme mesurer la quantité des superpositions existant entre les courbes de fréquence relatives au plus grand nombre possible de caractères.
Exact. Pourvu qu’on prenne soin, naturellement, de n’utiliser que les caractères biologiques significatifs et d’écarter ceux qui ne le sont pas. De toute manière, la science statistique moderne est parfaitement capable de nous fournir des techniques au point afin de déterminer la quantité des «superpositions».
Je n’ai donné là qu’un exemple de la façon dont le Systema Naturae peut être représenté avec objectivité : une hiérarchie de «cercle de formes» définissables en première approximation comme des nuages de probabilités insérés les uns dans les autres, les plus bas correspondant aux espèces et aux genres, les intermédiaires aux familles et aux ordres, et les plus élevés aux classes et aux types des catégories systématiques classiques. Mais en réalité, la tâche est plus ardue qu’il n’y paraît, car il faudrait évaluer, outre les relations morphologiques purement spatiales, les relations morphologico-temporelles.
|
Fig 9 La probabilité d’interaction, ou d’adhérence typologique, entre les caractères manifestés par les différents systèmes biologiques, peut être déterminée en évaluant l’aire de transvariation produite par la superposition de leurs courbes de variabilité (F, valeurs de fréquence ; Xi, valeurs d’intensité ; a, transvariation tendant vers zéro ; b, transvariation incomplète ; c, transvariation complète). L’adhérence typologique sera comprise entre les valeurs de 1 (transvariation complète) et 0, sans cependant jamais atteindre ce dernier absolument à cause de la nature asymptotique bien connue des courbes de variabilité (transvariation tendant vers zéro). Ceci permet aussi de comprendre pourquoi, dans la nature, aucune distinction ne saurait avoir un caractère absolu. En d’autres termes : les distinctions existent incontestablement, mais seulement dans un sens relatif, en tant qu’elles ne sont reconnaissables que dans le cadre de la continuité unitaire du Systema Naturae. Continuité et discontinuité sont donc complémentaires, l’une n’excluant pas l’autre. Les célèbres phrases de Linné, Natura non facit saltus et Species tot sunt diversae, quod diversas formas ab initio infinitum Ens creavit, restent vraies l’une et l’autre. |
Revenons à votre conception unitaire du monde vivant.
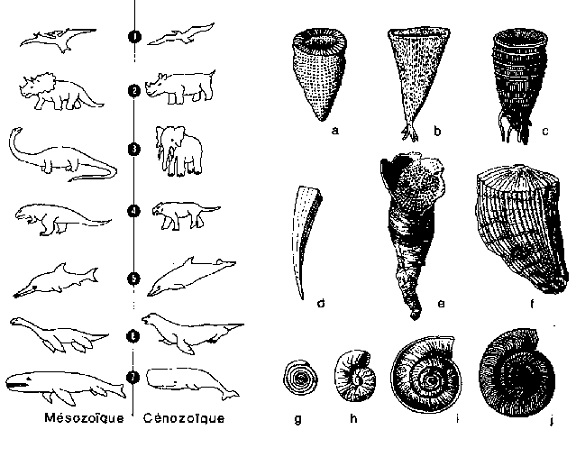
Il me semble que celle-ci, entre autres choses, commence aussi à faire deviner la possibilité d’expliquer les très nombreux cas de parallélisme morphologique existant entre des organismes souvent très éloignés entre eux dans le temps et dans l’échelle systématique : des cas qui, dans une optique simplement déterministe et phylogénétique, restent vraiment inexplicables. Pensez, par exemple, à la ressemblance entre la forme corporelle des Squales (poissons apparus au paléozoïque), des Ichtyosaures (reptiles marins du mésozoïque) et des Dauphins (mammifères marins apparus au cénozoïque), ou bien à la forme caractéristique de cône renversé et fixé au substrat, forme qui est commune aux Archéocyathidés, Spongiaires, Coelentérés, Brachiopodes (ex.: Richthofenia) et Mollusques (ex. : Hippurites) (figure 10). On est en droit de penser que des parallélismes de ce genre doivent obéir à quelque loi structurelle globale du système biologique naturel dans son ensemble chronotopique, et qu’ils peuvent donc être interprétés comme de véritables homologies inhérentes au système lui-même.
|
Figure 10 Quelques exemples de parallélisme morphologique entre Invertébrés (à droite) et Vertébrés (à gauche) ayant vécu à des périodes géologiques différentes. Invertébrés à la forme de cône renversé : a, archéocyathidé du paléozoïque ; b, éponge du mésozoïque (Ventriculites) ; c, corail du paléozoïque (Omphyma) ; d, conulaire du paléozoïque ; e, brachiopode du paléozoïque (Richthofenia) : f, mollusque bivalve du mésozoïque (Hippurites). Invertébrés à la forme de spirale plane : g, protozoaire foraminifère (Spirillina) ; h, annélide du paléozoïque (Spirorbis) ; i, mollusque gastéropode du mésozoïque (Discohelix) ; j, mollusque céphalopode du paléozoïque (Clymenia). Correspondances morphologiques entre hétérothermes (reptiles) du mésozoïque et homéothermes (oiseaux et mammifères) du cénozoïque : 1, Ptérosauriens et Oiseaux ; 2, Cératopsidés et Rhinocérotoïdés ; 3, Sauropsidés et Proboscidiens ; 4, Carnosauriens et Félidés ; 5, Ichthyosauriens et Dauphins ; 6, Plésiosauriens et Pinnipèdes ; 7, Pliosauriens et Baleines. |
Il est clair que les «cercles de formes» situés dans le futur restent inaccessibles à toute recherche scientifique. Mais il n’est pas exclu qu’on puisse arriver dès maintenant à en définir certaines caractéristiques fondamentales, à partir de règles fournies par une connaissance plus approfondie des «cercles de formes» des ères passées et de l’ère actuelle. En ce sens, il ne me semble pas que des thèses comme celles du paléontologue Dougall Dixon [17] doivent être considérées a priori comme tout à fait fantaisistes.
En fonction de toutes les argumentations précédentes, il faut alors conclure qu’une explication scientifique correcte des formes vivantes ne peut consister à tenter de les réduire à de simples effets de chaînes phylogénétiques de type déterministe et linéaire, les isolant ainsi du reste du système biologique unitaire dont elles sont des parties intégrantes et inséparables ; elle doit les ramener au système en en précisant, de la façon la plus objective possible, la place respective par rapport à toutes les autres formes composant et intégrant le système, dans sa totalité spatio-temporelle.
Savez-vous à qui je pense après avoir entendu vos conclusions ? À Carl Linné.
Certes. D’ailleurs, ne vous avais-je pas déjà précisé que mes opinions sont fort peu compatibles avec le vieil évolutionnisme «finaliste» ?
Par souci de clarté, je voudrais résumer brièvement la pensée de Linné en rapportant ce qu’écrit à ce sujet l’historien des sciences John Greene. Pour Linné, rappelle Greene, «l’histoire naturelle consistait à décrire les différentes productions de la Terre, leur apparition, leur formation, leurs relations de l’une à l’autre, et leurs mœurs. Comme Adam, le naturaliste commençait par donner un nom aux choses. L’âme de la science est la méthode, disait Linné, et la méthode consistait à assigner à chaque corps naturel un nom, en exprimant par là sa relation avec les autres corps. Les noms pouvaient être artificiels et arbitraires, dictés par de simples raisons de commodité, ou bien ils pouvaient être naturels, et exprimer par conséquent des affinités réelles entre les différents objets. La découverte de la méthode naturelle de classification, tel était le grand objectif du naturaliste (…) Durant toute sa vie, Linné ne renonça jamais à la recherche d’une méthode naturelle de classification, et ne se lassa jamais de proclamer que c’était là le but de la science» [18]. J’ai fait ressortir quelques phrases, qui me semblent particulièrement significatives et proches de votre façon de penser.
Oui. Chercher les relations réelles existant entre tous les vivants. En se fixant ce but, Linné affirmait ouvertement son adhésion à l’organicisme d’origine aristotélicienne. Mais il était par là inévitable que le naturaliste suédois, et après lui Georges Cuvier, qui en partageait les idées, finissent par attribuer au dessein naturel sous-jacent une stabilité absolue, appropriée à son comportement non purement mécanique. Et c’est précisément en ce sens que, comme eux, je n’hésite pas à me déclarer fixiste. La célèbre phrase de Linné, species tot sunt diversae, quot diversas formas ab initio Infinitum Ens creavit, reste à mon avis valable en règle générale. Disons que pour l’accorder à notre époque et pour lui donner une allure plus scientifique, je la transformerais ainsi : Taxa tot sunt diversa, quot diversae formae in spatii temporisque perpetuitate omnium animalium et satorum naturae congruenter inter se concinunt. C’est-à-dire : «Il existe autant de taxa différents que de formes qui interagissent harmonieusement entre eux, en conformité avec les lois de la nature, dans la continuité spatio-temporelle de tous les vivants» [19].
Y a-t-il actuellement d’autres hommes de science dont la pensée coïncide avec la vôtre ou s’en rapproche beaucoup ?
Je citerai avant tout Sir Fred Hoyle, le célèbre astronome britannique. Faisant preuve d’une audace et d’une ouverture d’esprit vraiment étonnantes, il aborde en effet et cherche à surmonter tous les points faibles et problématiques du paradigme évolutionniste dans une perspective non plus simplement biologique, mais physique et cosmologique. La solution proposée par ce vieux professeur est le résultat d’une longue réflexion, entamée avec Lifecloud [20] Diseases from Space et Evolution from Space, écrits avec son jeune collaborateur indien Chandra Wickramasinghe, et qui a trouvé son point culminant dans un magnifique ouvrage intitulé The Intelligent Universe [21]. J’ai trouvé dans ce livre des idées qui correspondent remarquablement à celles qui mûrissent en moi depuis des années. Les points de départ et d’arrivée de Hoyle — la critique radicale de l’«Évangile selon Darwin» et l’idée d’un univers «riche d’information» et «contrôlé de façon intelligente», «dans le sens inverse du temps, du futur vers le passé» — sont en effet identiques aux miens. Le reste, en revanche, me laisse assez perplexe, bien qu’on ne puisse pas du tout exclure qu’à la lumière des recherches qui seront certainement faites dans les prochaines années, cela finisse par s’avérer, pour une bonne part, exact.
Hoyle rejette nettement les assertions communes sur le problème de l’origine de la vie. «Toute la structure de la biologie orthodoxe — écrit-il — continue à soutenir que la vie est apparue par hasard. Mais plus s’accumulent les découvertes des biochimistes sur la grandiose complexité de la vie, et plus il devient évident que les possibilités d’expliquer l’apparition de la vie par le hasard sont si minces qu’elles peuvent être complètement écartées» [22]. La vie ne peut pas non plus être apparue sur la Terre à partir de la matière non vivante, mais doit être considérée comme «un phénomène cosmologique, peut-être l’aspect le plus fondamental de l’Univers» [23]. On a de bonnes raisons de penser que ce phénomène, comme tel et en tant qu’aspect fondamental de la substance même du monde physique, a toujours existé et est destiné à exister pour toujours.
Hoyle et Wickramasinghe soutiennent que, pour le moment, rien n’empêche — contrairement à ce qu’on croit généralement — de supposer que tout l’univers pullule de vie sous la forme de micro-organismes (bactéries, virus et viroïdes). La «poussière interstellaire», dont on a découvert au cours des dernières décennies qu’elle était répandue dans tout le cosmos, révèle en effet à l’analyse spectographique la même composition quantitative et qualitative que celle qu’on s’attendrait à voir dans une structure biologique. Il en va de même de l’analyse des atmosphères de Vénus, de Jupiter et de Saturne, ainsi que de la matière qui s’évapore des comètes lorsque celles-ci se rapprochent du Soleil. Si cette hypothèse devait s’avérer juste, il deviendrait alors très probable que les micro-organismes répandus dans l’espace parviennent à atteindre les planètes des différents systèmes stellaires à bord de comètes ou de météorites, y prospérant ensuite chaque fois que les conditions de la surface leur sont particulièrement favorables.
D’après Hoyle et Wickramasinghe, on peut alors penser que l’évolution de la vie sur notre planète s’est précisément produite à partir de microorganismes venus de l’espace et à travers une série considérable de mutations par «sauts» (ce qui s’accorde avec ce que présente la documentation paléontologique). Ces sauts auraient été provoqués par des infections successives de virus cosmiques, ou par d’autres micro-organismes, dont les gènes se seraient ajoutés au patrimoine génétique de précédentes formes de vie, le complexifiant au point de donner naissance à divers types biologiques, puis le modifiant continuellement au fil du temps. Hoyle et Wickramasinghe, en effet, estiment qu’au moins une bonne partie des épidémies brusques qui, depuis toujours et plus ou moins périodiquement, affligent l’humanité, sont justement produites par l’arrivée de micro-organismes venant de l’espace extérieur.
Cette idée de l’évolution par «infections successives venues de l’espace», bien qu’elle soit extrêmement séduisante, est peut-être la partie de la théorie en question qui laisse le plus perplexe. Mais on n’en a pas fini avec les surprises. Hoyle et Wickramasinghe (mais surtout Hoyle dans son dernier livre) affirment explicitement que l’évolution biologique, quand bien même se serait-elle produite comme le suggère leur hypothèse, ne saurait avoir eu lieu par hasard, mais doit avoir été constamment contrôlée par une «intelligence» agissant depuis le futur et au niveau cosmique. On retrouve ainsi une conception organiciste de la nature qui, pour l’essentiel, n’est pas différente de celle que je défends. Et il me paraît intéressant, à ce propos, de citer quelques phrases très significatives de Hoyle : «Cause et effet peuvent-ils opérer dans les deux sens du temps ? Les contradictions n’apparaissent-elles pas aussitôt dans ce système à deux voies ? Si nous continuions à penser séparément les deux sens du temps, la réponse serait oui et nous arriverions à d’insoutenables incohérences. Pour les éviter, les deux sens du temps doivent être reliés dans une espèce de circuit cohérent. Pour utiliser un langage précis, on devrait parler en termes de circuits, de tournants décisifs dans le temps, et non de cause et d’effet. Cause et effet ne deviennent une description convenable que dans des situations particulières qui impliquent des lieux de l’Univers et non l’Univers dans sa totalité (…) Ces considérations marquent un grand pas en avant vers l’élucidation d’un aspect tout à fait insuffisant dans la commune façon d’envisager les choses, selon laquelle le seul but du présent semble être d’engendrer le futur. Lorsqu’on atteint le futur promis, on tend vers un futur encore plus éloigné et ainsi de suite, à l’infini. On n’atteint jamais quelque chose de définitif, parce que chaque chose est abandonnée au moment où elle arrive. Toutefois, lorsque l’Univers est vu comme un circuit absolument fermé sur soi-même, rien ne peut être abandonné et chaque chose existe en vertu de tout le reste» [24].
Ce sont surtout pour ces conclusions, qui me semblent très bien s’accorder avec les miennes, que je crois que Hoyle et Wickramasinghe sont des scientifiques d’une importance vraiment centrale dans le débat actuel sur les explications ultimes du «phénomène vivant». Des hommes assurément plus importants que la figure, toujours affairée et prétentieuse, d’un Stephen Jay Gould, néanmoins considéré et présenté, sur le plan public comme sur le plan scientifique, comme l’un des esprits les plus profonds et les plus lucides de la pensée biologique actuelle.
«Nos opinions sur la biologie, ont écrit Hoyle et Wickramasinghe, ont été traitées de façon curieuse. Alors que nos recherches, particulièrement dans le domaine de la médecine, ont toujours été accueillies avec la plus grande courtoisie, personne ne s’est adressé à nous [en ce qui concerne la biologie] avec une indulgente sévérité, en nous donnant une petite tape sur la tête et en nous conseillant d’arrêter de dire des bêtises. Nous n’avons même pas été attaqués avec la dureté que nous attendions. Nos écrits ont plutôt été accueillis par le silence le plus absolu. La raison tient probablement au fait que chaque biologiste de formation s’est rendu compte dès le départ que, tôt ou tard, le mot ‘fin’ se serait montré, et impliquer le finalisme est, aux yeux des biologistes, le plus grand péché scientifique, encore plus exécrable que le fait d’émettre des doutes sur la validité du darwinisme» [25].
Je crois que ces remarques s’adaptent également plutôt bien à Dopo Darwin, étant donné la façon dont l’ouvrage a été accueilli en Italie.
__________________________________________________________
1 Thomas Kuhn, La structure des révolutions scientifiques, Flammarion, 1983.
2 Cf. à ce sujet : Yves Christen, L’heure de la sociobiologie, Albin Michel, 1979.
3 Ludwig von Bertalanffy, Théorie générale des systèmes, Dunod, 2ème éd., 1980, pp. 26-28.
4 Fritjof Capra, Le temps du changement, Rocher, 1984, p. 248, p. 257, pp. 262-263 (souligné par nous).
5 Raymond Ruyer, La gnose de Princeton, Fayard, 1978.
6 David Foster, The Intelligent Universe, Abelard, London, 1975.
7 Gregory Bateson, La nature et la pensée, Seuil, 1984.
8 Fritjof Capra, Le temps du changement, op. cit., p. 274.
9 Arthur Koestler, Janus. Esquisse d’un système, Calmann-Lévy, 1979.
10 Op. cit.
11 Adolf Portmann, Biologische Fragmente zu einer Lehre vom Menschen, Schwabe, Basel, 1969.
12 Arnold Gehlen, Der Mensch. Seine Natur und seine Stellung in der Welt, Athenaion, Wiesbaden, 1978.
13 Fritjof Capra, Le temps du changement, op. cit., pp. 283-284, p. 286.
14 Op. cit., pp. 265-266.
15 Il est intéressant de remarquer que l’idée de déroulement d’un plan correspond parfaitement au sens étymologique originel du terme «évolution». En effet, avant d’être employé dans le domaine scientifique, ce terme était surtout utilisé dans le langage militaire, pour désigner les manœuvres destinées à modifier les positions des unités d’infanterie, de cavalerie ou de marine, selon des directives établies par les responsables du commandement et de l’organisation.
16 Beverly Halstead, The Evolution and Ecology of the Dinosaurs, Eurobook, London, 1975.
17 Dougall Dixon, After Man, St. Martin’s, New York, 1981.
18 John C. Greene, The Death of Adam, Iowa State University Press, Ames, 1959.
19 Je remercie cordialement M. Aldo Ciompi, qui fut mon professeur de latin au lycée «E.S. Piccolomini» de Sienne, d’avoir bien voulu traduire pour moi cette version «contemporaine» de la phrase célèbre de Linné.
20 Trad. fr.: Le nuage de la vie. Les origines de la vie dans l’univers, Albin Michel, 1980 (N.D.T.).
21 Fred Hoyle, The Intelligent Universe, Dorling Kindersley, London, 1983.
22 Op. cit. (pp. 11-12 de l’éd. ital.).
23 Op. cit. (p. 12 et p. 161 de l’éd. ital.).
24 Op. cit. (p. 246 et pp. 248-249 de l’éd. ital. ; souligné par nous).
25 Fred Hoyle et Chandra Wickramasinghe, Evolution from Space, Dent and Sons, London, 1981 (p. 40 de l’éd. ital.).